CONVERGENCIA, Y Miembro de la Afiliación Científica Americana Miembro de la Asociación Americana de Antropología Miembro del Real Instituto de Antropología
Brockville, 1970 / Rev. 1977 Traducción del inglés: Santiago Escuain
|
 |
Índice
Acceso al artículo original -Convergence and the Origin of Man |
El
significado de la convergencia
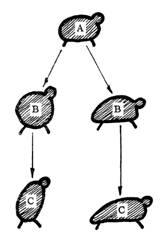
A CONVERGENCIA ES un fenómeno natural que según algunas de las autoridades más prestigiosas se da en todos los seres vivientes, plantas o animales. Es exactamente lo contrario a la divergencia, que en realidad es otro nombre para la evolución. Por convergencia se significa la tendencia que se observa en formas vivas no relacionadas filogenéticamente entre sí a responder a similares contingencias de la vida desarrollando estructuras semejantes. Estas «estructuras» incluyen no solo rasgos del esqueleto mismo sino órganos internos, órganos de los sentidos, fluidos corporales e incluso (al menos en las aves) rasgos como llamadas, coloración y hábitos de construcción de nidos. Es como si hubiera en la Naturaleza algún mecanismo intrínseco por el que cualquier animal o planta, al hacer frente a un problema que se deba resolver a fin de sobrevivir, pueda desarrollar una estructura, usando esta palabra en el sentido amplio anteriormente indicado, que resuelva el problema de la forma más económica y eficiente. Pero podemos ir más allá de esto y decir que estas soluciones exhiben una notable tendencia a conformarse tan estrechamente a un patrón, dependiendo de la naturaleza del problema, que tipos de animales sumamente diferentes (placentarios y marsupiales, por ejemplo), que no tienen una relación lineal por lo que atañe al pensamiento evolucionista actual, se desarrollan por separado en líneas tan similares que, si no tuviésemos otra información en sentido contrario, nos llevarían a la errónea suposición de que están muy estrechamente relacionados. Los dos diagramas en la Fig. 1 ilustran la diferencia fundamental entre divergencia y convergencia.
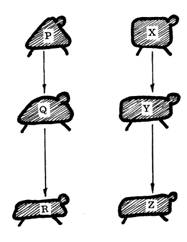
Fig. 1. Estos dos diagramas ilustran la diferencia fundamental entre divergencia y convergencia. El primer término es simplemente un sinónimo de evolución y supone que a partir de una sola forma animal A se puede observar que en las sucesivas generaciones los descendientes van difiriendo algo hasta que resultan en las formas B y C, de forma muy diferente. En cambio, en la convergencia, después de un período de tiempo se observará que los descendientes de unas formas P y Z, diferentes y no relacionadas, han llegado a parecerse mucho, de modo que se supondría que R y Z proceden de un antecesor común ... cuando de hecho no es así.
Debido al gran énfasis que los evolucionistas ponen en la importancia de la semejanza estructural (morfología) para establecer líneas de descendencia, el hecho de que puedan surgir semejanzas por medios totalmente no evolutivos constituye un serio reto para la actual teoría porque, por la misma naturaleza del caso, los evolucionistas no tienen ninguna otra forma convincente para construir sus «árboles» que por el estudio de la morfología (al menos, por lo que atañe al registro fósil). De forma que el reto de la convergencia constituye un problema fundamental. Como consecuencia, a pesar de los grandes esfuerzos dedicados a la investigación del hecho mismo, en años recientes se ha minimizado el fenómeno de la convergencia. De hecho, en muchos libros de texto se pasa totalmente por alto. En la edición de 1950 de la Chambers' Encyclopedia, aunque se trata la evolución con la extensión acostumbrada, la convergencia biológica ni siquiera se menciona en el índice general. El Oxford English Dictionary, de trece volúmenes, no se refiere al término «convergencia» con un sentido biológico en la lengua inglesa. En la edición de 1964 de la Encyclopedia Britannica, que ofrece páginas y páginas de texto sobre la teoría de la evolución, la convergencia se trata (bajo morfología) con un solo párrafo de dieciocho líneas, de las que una tercera parte se dedican en realidad al tema de la divergencia, que, como el autor observa, es simplemente otra palabra para la evolución.[1] Se nos dice que la convergencia no solo es infrecuente, sino que involucra solo semejanzas superficiales, mientras que de la divergencia se dice, naturalmente, que predomina en la Naturaleza. Pero, como se podrá ver con facilidad, lo cierto es precisamente lo contrario. Por otra parte, la convergencia, como ha dicho una autoridad evolucionista bien conocida, es un fenómeno casi universal. Este es un hecho constatado. Hay animales de los que se puede demostrar que no están relacionados y que han desarrollado estructuras o mecanismos precisamente similares y que son tan complejos que es totalmente inconcebible que puedan haber surgido puramente por accidente. Y estas convergencias no son en absoluto superficiales. Con frecuencia son de tal naturaleza que involucran a todo el animal. Por otra parte, la divergencia (o evolución) no es en ningún sentido un hecho demostrable. Si por «evolución» nos referimos meramente a las variaciones observables entre animales que se sabe que pertenecen a la misma especie (variedades de perros, por ejemplo) y que bajo ciertas condiciones siguen capaces de cruzarse y producir descendencia fértil, entonces estamos hablando de un hecho. Pero esta clase de evolución no arroja luz sobre el origen de las especies en el sentido más amplio. Hasta este punto, en tanto que la convergencia es un hecho establecido, la evolución es meramente una «teoría atractiva». Probablemente no hay ninguna teoría que haya sido promocionada con tanto fervor, defendida tan dogmáticamente y recibida con un reconocimiento tan universal sobre unas bases tan endebles como la teoría de la evolución. Es de sospechar que las debilidades inherentes en la actual teoría, que ya se están admitiendo en muchos lugares, explican la creciente hostilidad por parte de sus principales proponentes contra cualquier intento serio que se realice para reexaminar sus presuposiciones básicas. En una de sus últimas obras, G. G. Simpson,[2] como un niño silbando en la oscuridad, encontró necesario asegurar a sus lectores vez tras vez que la evolución es cierta; más aún, que es incuestionable. En This View of Life, Simpson repite ad nauseam sus «variaciones sobre el tema» en las páginas vii, 10, 12, 40, 51, 62, 63, 151, ¡y cinco veces en la página 193 en el espacio de catorce líneas! Hace algunos años, Ralph Gerard hizo esta significativa declaración, que es particularmente apropiada en el presente contexto:[3] «Cuando nos encontramos que mantenemos una opinión que la base en que se fundamenta suscita una especie de sentimiento de que indagar acerca la misma sería absurdo, evidentemente innecesario, improductivo, malo o incluso perverso, podemos considerar que se trata de una opinión no racional, y por ello probablemente fundada sobre evidencias inadecuadas.» Es característico de hasta qué punto el público ha sido extraviado por los evolucionistas que los bibliotecarios tienen el hábito de archivar los libros sobre evolución bajo el encabezamiento de «Ciencia», «Biología» o algo parecido, mientras que los libros que hacen un serio esfuerzo por exponer las falacias y debilidades del actual pensamiento evolucionista suelen ser archivados bajo el encabezamiento de «Religión». Existe un amplio consenso entre los investigadores acerca de que la singularidad del método científico reside fundamentalmente en investigar en busca del error más que de la verdad.[4] Por lo general, hace veinticinco años se nos hacía creer en la universidad que el método apropiado de investigación era primero acumular todos los datos posibles, y que una vez se había hecho esto, la verdad se haría evidente por sí misma.[5] Por medio de esto se suponía que se podía introducir una objetividad estricta. Nadie podría poner en tela de juicio la verdad a la que se hubiera llegado de esta manera. Lo que desde entonces se ha ido haciendo más evidente es que no hay ningún científico que actúe jamás de esta forma. No existe una mera «recolección de datos». Contemplamos la Naturaleza con anteojeras, conducidos por preconcepciones acerca de lo que esperamos encontrar. Y nuestra extracción de los datos es, nos guste o no, siempre un proceso selectivo; acabamos con capta («cosas tomadas»), no con data («cosas dadas»). No existen las «cosas dadas» —o bien sería más veraz decir, todo es «dado», y por consiguiente nos vemos obligados a seleccionar, porque no somos capaces de ver el todo. De modo que el método científico, tal como lo entendemos ahora, es no el de actuar como una especie de caja vacía adentro de la que invitamos a la Naturaleza a derramar su sustancia, sino el de actuar como filtro. Este filtro está estructurado por nuestras preconcepciones, nuestros prejuicios, por la actitud de nuestras mentes que, de hecho, es la fuerza motivadora que nos da la energía para investigar en primer lugar. La Naturaleza aborrece el vacío y no informará con utilidad a una mente que sea ella misma un vacío. De modo que la evidencia que encontramos en la Naturaleza es siempre «en favor» o «en contra» de alguna idea determinada que de manera consciente o inconsciente nos impulsa a realizar la búsqueda.[6] Entonces, si la investigación es un proceso con tanta carga de prejuicio, ¿cómo se puede conseguir la objetividad en absoluto? El dispositivo incorporado de seguridad que forma una parte esencial del método científico es la determinación de, si es en absoluto posible, descubrir el error en una hipótesis. Presuponemos que sabemos cuál es la «verdad», y luego investigamos con toda la sinceridad y honradez de propósito de que somos capaces para encontrar pruebas en contra. En este sentido, el método científico es una búsqueda en pos del error, no en pos de la verdad. Y cualquier hipótesis que no aliente a sus proponentes a realizar esta sincera búsqueda del error tiene que clasificarse como filosófica, no como científica. El agrio rechazo de parte del Establecimiento evolucionista a alentar a sus miembros a desafiar sus propias presuposiciones lo descalifica como empresa científica válida. Esta es la razón de que la convergencia sea una cuestión tan descuidada. Porque la convergencia presenta un desafío en dos aspectos. Por una parte es hostil, como ya hemos observado, a la actual dependencia en la morfología por parte de los paleontólogos para dibujar líneas evolutivas de descendencia en forma de árboles filogenéticos, sin los que ningún libro sobre este tema se consideraría bien presentado. Y es hostil, por otra parte, al actual rechazo que sienten la mayoría de naturalistas hacia la menor admisión de cualquier clase de vitalismo en los organismos vivos, que alentarían a la creencia de que la Naturaleza «sabe a donde va» respecto a su propósito. Que unas formas carentes de relación adopten paralelismos estructurales cuando se ven forzadas a resolver un problema similar en su ambiente implica que el proceso de cambio no es azaroso, debido al juego de la selección natural sobre mutaciones aleatorias, sino que está gobernado de alguna forma muy precisa por un mecanismo incorporado que no es meramente oportunista (por emplear el término predilecto de Simpson) sino que está claramente lleno de propósito. Y es que se ha de evitar a toda costa la idea de una conducta con propósito, en el sentido en que la contemplan los vitalistas, porque un propósito sugiere a Alguien con intención y con ello somos introducidos en el acto a la posibilidad de fuerzas que actúen independientemente o fuera de un marco estrictamente causal físico y químico. Esta Fuerza está totalmente más allá del alcance de la ciencia y por ello plantea un desafío a la reclamación implícita de omnicompetencia que hace la ciencia. Cuando los filósofos evolucionistas comenzaron a comprender las implicaciones de la convergencia, se la arrinconó calladamente como materia de investigación y discusión, incluso a pesar de que había recibido un amplio reconocimiento cuando, al principio, comenzó a acumularse una abundante prueba de la misma. Si este hecho hubiera sido mejor conocido antes de la publicación de El Origen de las Especies de Darwin, el curso de los acontecimientos en las ciencias de la vida pudiera haber tenido un giro muy distinto. Antes de 1858, cuando apareció por vez primera El Origen de las Especies, se había prestado mucha atención a la relación entre forma y función. Era la gloria de muchos de los científicos naturales de mediados del siglo diecinueve haber llevado hasta tal punto sus estudios por estas líneas que eran realmente capaces de reconstruir con notable precisión animales enteros en base a solo unos pocos huesos, sencillamente porque comprendían muy claramente que la forma está estrechamente relacionada con la función, de modo que si conocían la función que realizaba una estructura, podían recrearla en su totalidad en base a un fragmento relativamente pequeño. Virchow era un maestro en esto. Su influencia sobrevivió al surgimiento del darwinismo en personas como Wood Jones, que continuaron la tradición del estudio de la naturaleza en este sentido. Pero sus trabajos fueron pasados por alto por el Establecimiento que quedó crecientemente influido por la obsesión de Darwin con la morfología como la clave de las relaciones evolutivas. Es una feliz circunstancia que algunas de estas obras antiguas, que planteaban un desafío a las premisas básicas sobre las que se edificó la teoría de la evolución, han vuelto a aparecer en forma de reimpresiones. Así, se ha vuelto a publicar Mutual Aid, de Petr Kropotkin, donde se plantea un claro desafío al concepto de la Naturaleza enrojecida en colmillos y garras.[7] De mayor importancia en el contexto que nos ocupa, la obra de Leo Berg[8] Nomogenesis: or Evolution Determined by Law ha vuelto a salir a la luz como un renovado reto a estas suposiciones básicas, con su exposición de hasta que grado se encuentra en la Naturaleza el fenómeno de la convergencia a cada nivel de la vida y en el desarrollo de estructuras que son absolutamente esenciales para la continuidad del organismo. Berg es muy consciente de las implicaciones, y las resalta. En el siguiente capítulo examinaremos los hechos del caso tal como los han elucidado Berg y muchos otros. Y en el último capítulo veremos hasta qué punto la convergencia proporciona una explicación alternativa de los rasgos esqueletales de restos fósiles prehumanos y del primitivo hombre fósil, rasgos que se han presentado casi universalmente como prueba del origen animal del hombre.
Capítulo 2
El hecho de la convergencia
HA DE SER evidente que si la convergencia es en absoluto algo común, muchas formas que se han empleado para establecer árboles genealógicos con significación evolutiva puede que en realidad no constituyan árboles en modo alguno, sino, como lo expresó Manton,[9] «montones de ramitas» sin relación alguna en ningún sentido excepto en forma no evolutiva. Los árboles genealógicos son en tal caso totalmente ficticios, nada menos que tergiversaciones del curso de la historia biológica. No se puede poner mucha confianza en estas reconstrucciones filogenéticas. El mismo Darwin se sentía incómodo ante la significación de este hecho. En la sexta edición de El Origen de las Especies escribió:[10] «No se debería pasar por alto que ciertas variaciones fuertemente marcadas, que nadie clasificaría como meras diferencias individuales, aparecen con frecuencia debido a una organización similar sobre la que se opera, de lo que se podrían dar numerosos ejemplos con nuestra producción doméstica.» En 1876 escribió en una carta a Moritz Wagner:[11] «En mi opinión, el mayor error que he cometido ha sido no admitir suficientemente el peso de la acción directa del medio, esto es, la alimentación, el clima, etc., con independencia de la selección natural. ... Cuando escribí “El Origen” y durante algunos años después pude encontrar pocas evidencias de la acción directa del ambiente; ahora hay un gran cuerpo de evidencia.» Un curioso curso de acontecimientos siguió a la publicación de la obra de Darwin. Un resultado fue un creciente descuido del estudio de las relaciones entre forma y función, debido a una obsesión casi absoluta con el seguimiento de supuestas líneas de descendencia sobre la base de la forma. La morfología absorbió totalmente la atención de la mayoría de los estudiosos de las formas fósiles. El segundo resultado fue una investigación compensadora por parte de aquellos que se oponían al darwinismo (no necesariamente a la «evolución» como tal), en busca de ejemplos de paralelismo estructural entre formas vivientes que se hubieran desarrollado de forma completamente independiente y que no se considerasen como descendientes lineales. Con respecto a lo primero, Sir James Gray, el autor de una obra clásica sobre locomoción animal, escribió acerca de la teoría de la selección natural:[12] «Por extraño que parezca, un efecto inmediato de El Origen fue una marcada recesión en el estudio de la función animal. Hubo y sigue habiendo entre los estudiosos de la morfología una tendencia a atribuir a los órganos y estructuras una significación funcional para la que había o hay poca evidencia observacional. A este respecto, Evolution in Action, de Julian Huxley, no carece de culpa, por cuanto va mucho más allá de los hechos fisiológicos. ¿Está verdaderamente seguro el doctor Huxley de que la pérdida de los dígitos laterales por parte de los antecesores del caballo les proporcionó “una ventaja adicional en velocidad”?» Esta misma obsesión por la forma con independencia de la función condujo a Haeckel a formular su bien conocida pero ahora totalmente desacreditada teoría de la recapitulación. Sir Gavin de Beer observó:[13] «Ha
costado
más tiempo refutar la suposición de que las etapas de
desarrollo de un descendiente representan los tipos
ancestrales adultos debido a la forma superficial en
que se han presentado observaciones no cruciales como
pruebas en su favor. ... En muchos casos se puede
demostrar que la historia del desarrollo no puede
representar la historia filética, por la razón de que
si el antecesor adulto se pareciera al embrión
moderno, no hubiera podido ser funcional ... En otras palabras: se suponía y se sigue suponiendo que las semejanzas reales o superficiales constituyen prueba de descendencia lineal o de una estrecha relación genealógica, cuando puede tratarse de paralelismos funcionalmente determinados que resultan de una respuesta similar de organismos vivos a estímulos similares. Y de Beer concluía con una cita tomada del gran embriólogo Wilhelm His:[14] «Esta oposición a la aplicación de los principios fundamentales de la ciencia a las cuestiones embriológicas sería difícilmente comprensible si no procediera de un trasfondo de dogmatismo. No se admite ninguna otra explicación de las formas vivientes aparte de la herencia, y cualquier explicación que pueda basarse en otra debe ser rechazada. La moda actual exige que incluso la investigación más insignificante y neutral se presente vestida de una seda filogenética.» El doctor His escribió las precedentes líneas en 1888. Algunos años después, y tocante a conclusiones expuestas por antropólogos físicos, Wilson Wallis escribía con tono de decepción:[15] «Desde los tiempos de Darwin, la idea evolucionista ha predominado las ambiciones y ha determinado los hallazgos de los antropólogos físicos, a veces para detrimento de la verdad.» Y no cabe duda alguna acerca de que el célebre fraude de Piltdown nunca hubiera podido tener el gran éxito que tuvo excepto por el hecho de que Dawson proporcionó a los expertos precisamente lo que algunos de ellos creían que necesitaban. El Hombre de Piltdown era precisamente lo que los académicos habían reclamado. V. F. Calverton, en su introducción a The Making of Man, escribió:[16] «La simultaneidad misma con la que Darwin y Wallace dieron con la teoría de la Selección Natural y la supervivencia de los más aptos es una prueba manifiesta de la intensa actividad de esta idea en aquel tiempo. Todas las fuerzas en el ambiente económico y social jugaron su papel en el triunfo de esta doctrina.» En este mismo sentido se manifiesta A. K. Kroeber:[17] «Es evidente que hubo una determinada concatenación histórica en el pensamiento del mundo que posibilitó que el descubrimiento de Darwin desencadenara unas consecuencias de tal magnitud.» Como protesta contra este consenso de aprobación de una teoría que estaba sorprendentemente adaptada al Zeitgeist (como lo han observado muchos historiadores, entre ellos algunos de recientes), varias mentes independientes emprendieron una reevaluación de la evidencia. El príncipe Kropotkin reexaminó la comunidad de vida salvaje para ver si realmente había una «lucha por la existencia» y si solo los más aptos prevalecían. Descubrió en la Naturaleza una dinámica muy diferente, y expuso sus descubrimientos en su obra Mutual Aid. Hasta tiempos relativamente recientes este libro estuvo inaccesible. Pero el clima de opinión parece estar cambiando, y existe ahora una nueva demanda que ha justificado su reimpresión. De manera similar, en 1922 Leo Berg escribió su voluminosa y erudita Nomogenesis como protesta contra el interés desmesurado por la morfología con exclusión de la función. En la actualidad quizá hubiera titulado su obra Convergencia, porque es de esto de lo que trata. Esta obra también se ha vuelto a publicar, y, cosa sorprendente, de mano del Instituto de Tecnología de Massachusetts. Poco después de que apareciera esta primera edición, en 1935 Sir Wilfrid LeGros Clark estaba dispuesto a admitir:[18] «En la evaluación de las afinidades genéticas, las diferencias anatómicas son más importantes como evidencia negativa que las semejanzas anatómicas lo son como evidencia positiva. Se hace evidente que si esta tesis se lleva a su conclusión lógica, será necesario demandar un ámbito mucho más amplio para los fenómenos del paralelismo o de la convergencia en evolución de lo que los evolucionistas han concedido en general. La realidad es que las minuciosas y detalladas investigaciones que han realizado anatomistas comparativos en años recientes han puesto de relieve que los paralelismos en el desarrollo ha tenido lugar a gran escala y que ya no deben considerarse como una curiosidad incidental que haya tenido lugar de forma esporádica en el curso de la evolución. Lo cierto es que apenas si es posible para los no anatomistas comparativos darse cuenta de lo fundamental del papel que este fenómeno ha tenido en el proceso evolutivo [¿de desarrollo? —inciso mío].» En cambio, en la actualidad se oye muy poco acerca de esta cuestión, como se ve por el silencio de las enciclopedias. El mismo Sir Wildfrid, en su contribución al coro darwinista en las publicaciones que aparecieron con motivo del Centenario de Darwin (1958), escribió:[19] «Se debería hacer referencia a los fenómenos evolutivos de la convergencia y del paralelismo, porque es bien sabido que dichos fenómenos pueden llevar a semejanzas estructurales que, si se toman por sí mismas, pueden conducir a error. El término “convergencia” se aplica a la tendencia ocasional de que tipos relacionados distantes se simulen entre sí respecto a proporciones generales o en el desarrollo de adaptaciones análogas como respuesta a unas necesidades funcionales similares.» ¿Qué sucedió durante estos aproximadamente 25 años para que los fenómenos «a gran escala» de los que Clark decía que «ya no deben considerarse como una curiosidad incidental» se hayan convertido en una «tendencia ocasional»? Quizá se había hecho crecientemente claro durante los años transcurridos que la admisión de la realidad de la convergencia a gran escala era sumamente contradictoria respecto a muchos de los árboles genealógicos comúnmente exhibidos y que pretendían exponer una descendencia evolutiva lineal basándose puramente en la morfología. Tan crucial es la morfología que el antropólogo Franz Weidenreich formuló el siguiente principio:[20] «Al proceder a la determinación del carácter de una forma fósil determinada y de su puesto especial en la línea de la evolución humana, solo se deberían tener en cuenta sus rasgos morfológicos como base decisoria: ni la situación del emplazamiento de donde fue recuperado, ni la naturaleza geológica del yacimiento en el que estaba sepultado tienen importancia.» Pero, ¿qué valor tiene esta exigencia de que se ignore la evidencia geológica y que solo se considere la apariencia física? A la luz de la posibilidad de que la estructura sea enteramente resultado de circunstancias ambientales o históricas y que no tenga nada que ver con la edad geológica, el argumento carece de toda validez. Sir Solly Zuckerman, aunque era un evolucionista plenamente convencido, admitió abiertamente lo siguiente:[21] «Diversas pautas genéticas pueden tener idénticos efectos fenotípicos (de modo que) cuando tratamos con material fósil limitado o relativamente limitado, la correspondencia en rasgos morfológicos simples o en grupos de caracteres no implica necesariamente identidad y relación filética.» Por la frase «diversas pautas genéticas», Zuckerman se está refiriendo al hecho bien reconocido de que donde las circunstancias «demandan» que un animal esté equipado con algún órgano particular (por ejemplo, una clase especial de ojo), que se dará la tendencia a que aparezca este órgano aunque el animal no comparta un conjunto genético que sea responsable del mismo órgano en alguna otra especie. Así, los conjuntos o pautas de genes que difieren pueden sin embargo llevar a la producción de estructuras similares en animales no relacionados. Wood Jones argumentó con energía que había alguna especie de «fuerza vital» en la naturaleza que tenía como resultado el surgimiento de toda clase de estructuras especializadas en animales que capacitaban a sus poseedores a afrontar las exigencias particulares de sus vidas. Estas estructuras, según era su convicción, podían aparecer «de la nada», por así decirlo, casi bajo pedido. Da muchos ejemplos en su clásico pequeño volumen Trends of Life, todos los cuales se escogieron para demostrar que en cierta manera misteriosa «la Naturaleza sabe adonde va». Al igual que Leo Berg, y ahora incluso más recientemente Sir Alister Hardy,[22] Jones también estaba persuadido de que había poco o ningún elemento de azar en este fenómeno. En sus palabras:[23] «Desde la aceptación de la teoría de la Evolución propuesta por Darwin se han realizado muchos intentos, por parte de distinguidos biólogos (como Gaskill y Patten), para demostrar que los invertebrados realmente “evolucionaron” transformándose en vertebrados; pero toda la evidencia disponible deja muy claro que los dos grandes filums surgieron independientemente el uno del otro ...» Cuando los propagandistas dogmáticos de la teoría darwinista de la evolución le dijeron al gran público que algo tan complejo como un ojo había llegado a la existencia gracias a una vaga fuerza conocida como Selección Natural que había actuado sobre pequeños cambios estructurales producidos al azar, su credulidad fue llevada al extremo. Es posible que la fe del público se hubiera derrumbado por completo si les hubieran pedido que creyesen que este proceso mecanicista al azar había producido el ojo de los vertebrados y el de los invertebrados de modo independiente en ambos casos, y más que esto, que había permitido que los invertebrados adquiriesen al menos tres clases diferentes de ojos de modo independiente, dentro de los límites de su propio filum (los ojos de foco simple, bifocal, y compuesto). No solo los ojos, sino los oídos, corazones y las branquias, y todo lo demás, se han desarrollado dos veces de forma completamente independiente en los dos grandes filums. Además, como veremos, la lista que da Jones apenas si araña la superficie de los paralelismos menos evidentes existentes. Alguna ley hasta ahora no reconocida o mal definida ha estado operando y gobernando toda la vida. Naturalmente, en el actual clima de opinión, estos conceptos están demasiado saturados de metafísica para que el Establecimiento lo aliente. Se rechazan sin más miramientos. Todo ha de ser abandonado al azar. Evolución y Azar son conceptos virtualmente sinónimos, y quizá LeGros Clark (igual que otros) había comenzado a darse cuenta de que la convergencia favorecía el desarrollo por ley y no por azar, de una manera demasiado incisiva. Está muy claro que esta convicción impulsó a Leo Berg a escribir su estudio clásico sobre la convergencia y a darle un título más preciso: Nomogénesis: Evolución determinada por ley. Naturalmente, la idea de que pudiera haber alguna ley gobernando el desarrollo de las formas vivas a través de las edades no debería generar más temor que el concepto de ley en física. Pero los acontecimientos físicos del pasado no han evidenciado ningún progreso de simple a complejo, de inferior a superior, de mayor dependencia del medio ambiente a una menor dependencia del mismo, de carencia de un propósito consciente a un grado muy elevado de propósito, etcétera, al modo de los seres vivientes. En este sentido hay una dirección del desarrollo de la vida que no es evidente en el mero orden físico. Y la idea de «dirección» en conformidad a una «ley», y en una magnitud significativa en contra de la regla por otra parte universal de «degeneración» (entropía) suscita inevitablemente el espectro del propósito. Y un propósito implica a Alguien que lo tiene. Aquí es donde está el quid de la cuestión ... Berg vio que la convergencia era tan predominante que pudo escribir sin vacilación alguna: «La convergencia, y no la divergencia, es la regla, no la excepción. Esto parece ser predominante, tanto entre las plantas como entre los animales, presentes, recientes y extintos.»[24] Y en la última reimpresión de su obra le encontramos diciendo:[25] «A partir de los ejemplos expuestos en esta sección, es evidente que la convergencia afecta a los órganos más importantes y fundamentales para la existencia, y no meramente a los rasgos externos.» Y de nuevo, más adelante:[26] «A partir de los numerosos ejemplos que se han ofrecido en este capítulo, y su número se podría multiplicar fácilmente, hemos demostrado que la convergencia afecta a los órganos más fundamentales en los animales y en las plantas, que el fenómeno está ampliamente difundido, y que los puntos de semejanza que se han atribuido a la descendencia común se deben con frecuencia a la convergencia.» Antes de examinar las pruebas con más detalle, será oportuno observar, para ser justos con otros escritores posteriores, que aunque la convergencia es una doctrina demasiado peligrosa para que pueda recibir demasiado énfasis, está sin embargo admitida con bastante extensión por el Establecimiento. G. G. Simpson estuvo dispuesto a admitir:[27] «En
la
convergencia se da la misma clase de desarrollo
oportunista de un estilo de vida por parte de grupos
diferentes —tratándose en este caso de grupos
disimilares (o menos similares) en tipo adaptativo
para comenzar. La tendencia hacia una mayor semejanza
de adaptación involucra ... características
funcionales y estructurales convergentes. Los grupos
pueden estar casi relacionados o puede que estén
relacionados solo muy de lejos. ... Los insectos y las
aves están tan distantemente relacionados que apenas
si puede seguirse ninguna homología determinada entre
sus partes, y sin embargo convergen, a veces de manera
muy estrecha. Alfred S. Romer[29] observó que «el desarrollo de largos cuerpos fusiformes para soportar una cola dorsal se ha dado en al menos dos y quizá cinco líneas separadas». Y comunicó que Radfield le había sugerido que se trataba de mecanismos de regulación térmica. Esto me parece muy probable. Una estructura así se observa por ejemplo en el Dimetrodon. Boule y Vallois extendieron algo más este principio:[30] «Sería importante conocer si podemos aceptar o no la existencia de fenómenos de convergencia de caracteres bioquímicos, análogos a la convergencia de un orden morfológico. No hay razón por la que una evolución [¿desarrollo?] similar en dos grupos diferentes no debiera ir acompañada por una evolución en paralelo de fenómenos atribuibles a la bioquímica. Parece que los naturalistas no han prestado atención a este punto.» [El interrogante es del presente autor.] Como veremos, los bioquímicos como tales sí que han prestado atención a este punto, pero los evolucionistas dogmáticos no han prestado atención a los bioquímicos. Refiriéndose al linaje humano, Ruggles R. Gates escribió:[31] «La abundancia de tipos convergentes involucra también el reconocimiento de que grupos como los de los mamíferos, que se consideran ahora como uniformes [es decir, procedentes de un solo antecesor] han tenido un origen polifilético (esto es, independiente).» Alfred Romer[32] llegó tan lejos como para decir que «la presencia conocida de paralelismos en tantos casos, y la sospecha de su presencia en otros sugiere que la convergencia puede haber sido un fenómeno casi universal». Y Simpson parecía estar lanzando el mismo llamamiento a la cautela en contra de la suposición demasiado precipitada de una relación basada en homologías, al decir: «¡Sanger ha demostrado que la composición de la insulina de las ballenas de esperma es idéntica a la de los cerdos y bien diferente del de las ballenas sei! Es cierto que solo está implicada una secuencia de tres aminoácidos, y tanto las diferencias como las semejanzas podrían ser accidentales sin incluso una verdadera convergencia, pero ahí tenemos una lección.»[33] Para dar otro ejemplo, Herbert Friedman, Conservador de la División de Aves en el Museo Natural de los Estados Unidos, presentó, en un artículo acerca de las correspondencias en aves, un estudio de los extraordinarios paralelismos en especies no relacionadas de aves, incluyendo patrones de alimentación, llamadas y construcción de nidos, además de en coloración y detalles estructurales. Decía él:[34] «Cuanto
más
completo se hace nuestro conocimiento de cualquier
grupo determinado de organismos, tantos más casos
vienen a la memoria. ... La cantidad de ejemplos se
puede alargar hasta un extremo fatigoso. ... Llegamos hasta aquí, entonces, por lo que se refiere al reconocimiento del hecho en sí. Es a la obra de Leo Berg en particular a la que hemos de remitirnos para el examen más exhaustivo de la evidencia. Hasta cierto punto, tenemos una indicación del renovado interés en esta cuestión como un todo en el hecho de que su trabajo original, publicado por vez primera en ruso en 1922, y en una traducción inglesa en 1926, se ha puesto otra vez a disposición para su estudio en 1969. Consideremos unos pocos ejemplos de órganos y fluidos corporales. Según Berg, la placenta, mediante la que el embrión queda conectado con el cuerpo de la madre y a través de la cual recibe su alimentación y se libra de sus productos residuales, se ha formado de manera independiente en diversos grupos de animales, incluyendo los polizoos (un cierto tipo de parásito de los peces), el Penpatus (orugas no segmentadas exteriormente), ciertos insectos y escorpiones, los tunicados (mantas marinas), ciertos tiburones, ciertos marsupiales, y naturalmente todos los mamíferos placentarios. Berg observa:[35] «Todo en la anatomía, embriología y paleontología de los mamíferos nos inclina a compartir la opinión de Abel: los monotremas, los marsupiales y los placentarios son tres ramas paralelas que han surgido de forma independiente entre sí.» Berg señalaba que la clorofila y la hemoglobina son sustancias análogas, pero que han surgido de forma totalmente independientes como portadoras de vida:[36] «Manoilov[37] ha descubierto una reacción para distinguir la sangre del varón de la de la mujer; es digno de mención que esta misma reacción permitiera distinguir entre el sexo macho y hembra en plantas deciduas como el arce (Acer negundo), la ortiga (Lychnis dioica) y la Vallisneria.» Acerca de esto, Berg comenta:[38] «Un paralelismo fisiológico así indica que la elaboración de sustancias químicas (que en último término afectan a la morfología así como a la fisiología) queda sometida a ciertas leyes.» Mientras tratamos de la sangre, se puede observar de pasada que C. L. Prosser observó lo siguiente, en un artículo inspirado por las «celebraciones» del Centenario de Darwin:[39] «Las hemoglobinas, diferentes en la proteína pero semejantes en el grupo heme, han evolucionado muchas veces por separado —en los cordados, unos cuantos moluscos, algunos entomostreones, ciertos anélidos, numerosos holotúridos, unos pocos insectos dípteros, e incluso algunas nitrobacteriáceas.» Por otra parte, la sangre caliente ha aparecido naturalmente dos veces de forma independiente, en las aves y en los mamíferos. La transformación de un animal de sangre fría a otro de sangre caliente implica el surgimiento de unas estructuras de los sistemas central y periférico que son casi increíblemente complejas. Berg observó que en ciertas plantas insectívoras se ha desarrollado un agente fermentante semejante a la pepsina del sistema digestivo animal, para capacitarlas para usar alimentos proteínicos; este agente es segregado por un órgano homólogo.[40] Pero, cosa curiosa, la pepsina aparece solo en los animales superiores; su presencia en invertebrados sigue estando en duda. Berg se refirió al desarrollo de ojos bifocales estructurados de modo muy semejante en peces y en los escarabajos de Whirligig, carentes de toda relación entre sí.[41] Estos ojos bifocales permiten al escarabajo ver normalmente en el aire y a la vez estar vigilante debajo del agua, mientras que en ciertos peces se observa lo contrario. En ambos casos una banda horizontal divide el ojo en una sección superior y otra inferior, siendo la lente de la primera idónea para ver en el aire, y la de la segunda para ver en el agua. De modo que este órgano tan complejo ha surgido en dos ocasiones de forma totalmente independiente. Además, y tal como lo observó Rendle Short,[42] los ojos del pulpo son precisamente iguales a los de los mamíferos, incluyendo al hombre, y este paralelismo se extiende a la estructura de la córnea, del iris, del músculo ciliar y de sus procesos, y de la retina. Sin embargo, es evidente que no hay relación «evolutiva» entre estas dos clases de criaturas vivientes. Berg comenta:[43] «Nos encontramos con ojos con una lente de forma independiente en los gusanos anélidos, en artrópodos y cefalópodos. En este último caso nos encontramos con la retina, la córnea, el iris, el proceso ciliar e incluso (en algunos casos) con párpados.» Zawarzin[44] se refirió a los ojos como «un principio estructural relacionado con la facultad de la visión común a todo el mundo animal». Que una estructura así con todas sus partes componentes (suministro de sangre, glándulas lacrimales, mecanismos neuromusculares de control y las áreas visuales asociadas del cerebro) se haya formado tantas veces en unas criaturas tan diversas es algo que desde luego está fuera de la capacidad del puro azar actuando por medio de la selección natural sobre cambios aleatorios en el conjunto genético. Esto sí que es fe en milagros. Tocante a fenómenos tan extraños como la luminosidad o fosforescencia, Berg observó:[45] «Los órganos luminosos o fosforescentes, enigmáticos tanto en su función como en su origen, se desarrollaron de forma independiente en los grupos más diversos de peces marinos como los tiburones (Spinax y otros), en los stomiatidae, los scopalidae, en antennariidae, y otros.» Un arma tan notable como la capacidad de lanzar un choque eléctrico muy potente ha aparecido también de forma independiente en tres animales acuáticos: el anguila eléctrica (una especie de agua dulce de Norteamérica), el torpedo (de amplia distribución en los océanos) y el Malapterurus en África.[46] Volviendo a la cuestión de la defensa, las púas han surgido en animales por otra parte tan inofensivos como el hormiguero monotrema australiano (localmente conocido como «puercoespín»), el verdadero puercoespín roedor (Hystrix) común en Europa y África del Norte, el puercoespín sudamericano (Synetheres), también el erizo común, y el pequeño y punzante Ericulus de Madagascar.[47] En su libro Animal Weapons, que trata de la defensa en un momento de una total impotencia, Philip Street observó acerca de un caso de un desarrollo paralelo en insectos en la etapa larval:[48] «La evolución convergente, por la que dos tipos de animales carentes totalmente de relación entre sí evolucionan a estructuras semejantes para un propósito similar con total independencia entre ellos es un fenómeno sumamente interesante. Desde luego, no hay relación posible entre los diversos tipos de gusanos cilíndricos y las larvas de los frigánidos, y sin embargo estas larvas, comúnmente conocidas como tricópteros, construyen tubos para su propia protección que son notablemente similares a los producidos por los anélidos marinos.» Dejando ahora órganos o estructuras, podemos observar que lo mismo es aplicable a animales enteros. Refiriéndose a la fauna australiana, David Lack dijo:[49] «Australia fue colonizada por mamíferos marsupiales que, en ausencia de formas placentarias, evolucionaron a formas homólogas a los zorros, lobos, topos, ardillas, conejos, ratas, osos hormigueros y ardillas voladoras, que se parecen, a menudo mucho, a sus formas correspondientes entre los mamíferos placentarios de otros continentes.» Sin embargo, no están relacionadas entre sí. Las siguientes ilustraciones, tomadas de la obra de Sir Alister Hardy, The Living Stream, muestran la gran correspondencia en detalle estructural que pueden presentar estas formas paralelas.[50] La rata del desierto y los jerbos (véase Fig. 2) responden evidentemente a las presiones del medio ambiente al desarrollar la misma excepcional forma de conjunto que les posibilita moverse rápidamente en la arena suelta saltando como un canguro en lugar de correr. El cráneo del lobo de Tasmania no se puede distinguir del cráneo del lobo norteamericano o europeo (véase Fig. 3).[51] La gama de variabilidad es común a ambas formas. Aun más notable es la estrecha semejanza entre los topos placentarios y marsupiales (véase Fig. 4), que han desarrollado una configuración casi idéntica de pies excavadores, nariz y boca, estructura de los ojos, y aperturas de los oídos, que tienen el propósito de impedir que entren partículas por el orificio auditivo.[52] Pero estas dos criaturas no están relacionadas. G. G. Simpson[53] ha dado una ilustración del marco estructural que sustenta el ala de un murciélago y de una mosca, y cree que constituye otro ejemplo de desarrollo convergente para resolver un problema común de ingeniería. Wood Jones lo expresó de forma muy sucinta:[54] «Así, parece cosa cierta que unas estructuras que se han desarrollado para satisfacer estas necesidades comunes pueden exhibir un parecido considerable entre sí, aunque los animales que las manifiesten puedan carecer completamente de relación por parentesco o descendencia. Debido a que hay tantas necesidades basales comunes a todos los animales y que estas necesidades funcionales se satisfacen mediante el desarrollo de estructuras apropiadas, es de esperar que se pudiera detectar un plan de base común de partes y órganos. ...
Fig. 2. (A) El jerbo marsupial (Antechinomys laninger) y (B) el jerbo placentario (Dipus hirtipes), redibujados respectivamente de la obra de Troughton's Furred Animals of Australia y de Cambridge Natural History.
Fig. 3. El lobo marsupial de Tasmania (Thylacinus cynocephalus), con (A) su cráneo comparado con (B), el del lobo placentario (Canis lupus), dibujado a la misma escala, a partir de especímenes del Museo de la Universidad de Oxford.
Fig. 4. (A) El topo marsupial (Notoryctes), redibujado de Cambridge Natural History, y (B) el topo placentario (Talpa europea), dibujado a partir de una fotografía.
De modo que las homologías no se deben ni al azar ni a la descendencia, sino a un factor de diseño incorporado. Se podría argüir que no poseemos pruebas directas de que se puedan atribuir los cambios sustanciales en estructura a factores ambientales. Pero sí que tenemos tal prueba. Por ejemplo, el fenómeno de la carencia de cuernos en ganado que normalmente está dotado de cuernos se observa cuando se desplazan manadas a regiones en las que ya se conoce esta condición de carencia de cuernos. Este tipo de ganado se encuentra en Europa, África y América del Sur.[55] Asimismo, unas manadas suizas trasladadas a Hungría desarrollaron cuernos y patas de mayor longitud, como los ganados locales. En otro caso, ganados transportados desde los Alpes bávaros al estado de la corona de Altenburg en Hungría no solo desarrollaron los cuernos más largos comunes en aquella región, sino que sus cuernos se volvieron más duros y adquirieron una forma peculiar de lira característica de la variedad magiar local, y el cráneo se volvió más estrecho.[56] Estas diferencias estructurales forman con frecuencia la base sobre la que los taxonomistas modernos distinguen entre especies y géneros, e incluso entre órdenes más elevados de clasificación. Los investigadores creían que estos cambios se debían totalmente a los efectos del clima y de la naturaleza del terreno de Hungría. No se debieron a hibridación. Unos peces (Zoarees viviparus) transferidos del Fiordo Ise al Fiordo Roskilde en Dinamarca, a unos pocos grados de latitud, aumentaron la cantidad de vértebras, desde la media nativa de 108 a una media más elevada de 114,6 vértebras.[57] Estas líneas no se mezclaron con peces que ya pertenecían a esta zona. De modo similar, moluscos terrestres de diferentes géneros han desarrollado formas paralelas de conchas, marcas y patrones decorativos de forma totalmente independiente.[58] La lista se podría alargar hasta el aburrimiento. Pero el fenómeno funciona en ambos sentidos. Los desemejantes pueden volverse semejantes, y los semejantes pueden llegar a diferir —la relación real o su ausencia no se reflejan necesariamente en el resultado final. Esto es de aplicación también a los simios y a los hombres, aunque los datos reales que poseemos muestran más bien que el cráneo humano degenera más fácilmente hacia una forma simiesca que el caso contrario, de que un cráneo de simio se vuelva más humanoide. Si se ignora el tamaño, ciertos simios ahora extintos se desarrollaron durante un largo período de tiempo, según se dice, hasta una forma más humanoide incluyendo la adquisición de una postura parcialmente erguida, un desarrollo algo reducido de los arcos superciliares y un patrón de dentición más humanoide. Estos cambios son posibles y pueden haber aparecido debido a factores ambientales con influencias secundarias, que resultasen de cambios en hábitos alimenticios al modificar las condiciones del medio ambiente la fauna y flora locales que comprendían el suministro alimentario. En el siguiente capítulo examinaremos la extensión en que el cráneo humano pueda quedar estructuralmente «degradado» hasta que se parezca más de cerca en ciertas formas al cráneo símico. Se puede demostrar que factores como el clima, la dieta y ciertos hábitos culturales relacionados con comer (como, por ejemplo, el uso o no uso de cuchillos) realizan cambios en la morfología craneana que brutalizan los rasgos y que proporcionan alguna justificación, en ausencia de cualquier otro criterio conductor, para suponer que los hombres fósiles estaban filogenéticamente más cerca de los simios que del hombre moderno.
Capítulo 3
Las
implicaciones de la convergencia
ES COSA bien sabida que el cráneo humano es lo suficientemente plástico para que se pueda modificar, en la etapa adulta, hacia una forma más simiesca si el hombre simula los hábitos alimenticios del simio de alguna u otra manera. Esta cuestión es susceptible de examen debido a que muchos de los cráneos del hombre primitivo han quedado indudablemente deformados en dirección del cráneo símico debido a lo que Portmann[59] hubiera designado como razones «históricas» (en contraste a genéticas). Esta deformación puede darse dentro del transcurso de una sola vida. Evidentemente, la descendencia no la hereda, pero si estas condiciones de vida persisten durante varias generaciones, lo más probable es que quedarán algunos cráneos preservados en forma fósil cuya configuración podría dar la impresión de que sus poseedores no estaban muy alejados por descendencia de algún antecesor simiesco subhumano, cuando, en realidad, no es necesario suponer tal clase de relación. Hace años, Wilson D. Wallis[60] observó que «La
evidencia
de los restos humanos prehistóricos no justifica de
por sí la inferencia de una descendencia común con los
simios. Basamos esta conclusión en el hecho ... de que
prácticamente todos los cambios en la estructura
humana que se pueden seguir a través de los restos
prehistóricos son resultado de cambios en
alimentación y hábitos. Un volumen del Simposio de Ciba se dedicó a un estudio de la vida de los esquimales. Este tema es particularmente relevante en el presente contexto debido a que este extraordinario grupo humano es considerado a menudo como un «modelo» bastante preciso del hombre del Paleolítico. Escribiendo en este número, Erwin Ackerknecht observaba:[61] «Los huesos malares y las mandíbulas de los esquimales son enormes, quizá bajo la influencia de la intensiva masticación que tienen que practicar, lo que también resulta en un enorme desarrollo de los músculos de masticación. Los dientes de los esquimales quedan a menudo desgastados hasta las encías, como los dientes de los animales, debido a su empleo excesivo.» En la Fig. 5 vemos una cara esquimal masculina característica, con el bosquejo de la forma del cráneo que indica que el mayor grosor aparece en las quijadas y no en la región de las sienes. En cambio, la cabeza del Niño Azul de Gainsborough, en la Fig. 6, muestra cómo una dieta refinada tiende a producir una forma de cabeza de otra clase, con la mayor amplitud en la región temporal. Se ha observado también que el cráneo esquimal exhibe ocasionalmente una «cresta» a lo largo de la parte superior, lo que es el resultado directo de la necesidad de una unión o anclaje más fuerte para los músculos de la mandíbula que se usan de manera mucho más extensa. Esto se observará en la Fig. 5 (b), y se debería comparar con la cresta que se indica en los cráneos de los fósiles supuestamente humanos en la Fig. 7 (c, d, e). Queda muy claramente marcada en el caso del cráneo del gorila en la Fig. 7. William Howells hace este comentario:[62] «Los gorilas tienen una mandíbula muy pesada y poderosa, y los músculos que la cierran (que en el hombre forman una delgada capa sobre y alrededor de la sien, donde se pueden sentir cuando se mastica) son tan grandes que presentan un gran grosor en la parte superior de la cabeza, de alrededor de cinco centímetros de espesor, cubriendo prácticamente la voluminosa arcada ósea sobre los ojos que es tan prominente en el cráneo, y originando una cresta ósea en medio solo para separar y dar un punto de anclaje a los músculos de ambos lados.» Así, en el cráneo esquimal y en el del gorila aparece en ocasiones un cierto paralelismo que en modo alguno indica una relación genética. La explicación de la cresta esquimal es histórica (es decir, cultural). Respecto a esto, podemos volver a citar a Howells:[63] «La poderosa mandíbula de estos animales suscita, al masticar, una tremenda presión hacia arriba contra el rostro, y los arcos superciliares constituyen un fuerte límite superior que la absorbe.» Si se somete al hombre a alimento sin cocinar y se le obliga en ausencia de cuchillos a arrancar la carne del hueso, los músculos, en su desarrollo, encontrarán la forma de fortalecer su anclaje a lo largo de estas arcadas óseas. Además, si no aparece en la dieta un factor que endurezca el hueso en los primeros años de la vida cuando se encuentran estas tensiones al principio, es inevitable que el cráneo se deprima mientras está todavía en un estado relativamente plástico, y la parte frontal del cráneo será baja y en pendiente de modo que carecerá de la elevada bóveda que solemos asociar con el hombre culto. Así, las grandes arcadas superciliares del Sinanthropus, tan semejantes a las del Pithecanthropus, son, como observó Ales Hrdlicka hace algunos años, «un rasgo que se debe correlacionar con un poderoso mecanismo mandibular».[64]
Fig. 5. Compárese la forma de esta cabeza esquimal con la cabeza del «Niño Azul» en la Fig. 6. Este dibujo se basa en una foto reproducida en la cubierta del Simposium de Ciba (Vol. 10, nº 1) y mantiene las proporciones exactas: (A) un bosquejo simplificado; (B) un antiguo cráneo esquimal, que exhibe la cresta (ligeramente exagerada) en la parte superior y frontal de la cabeza.
Fig. 6. Esta cabeza se basa en el cuadro de Gainsborough «el Niño Azul», y está dibujada exactamente a escala. Muestra claramente la influencia de lo que podría designarse como una dieta culta. La parte ancha de la cabeza está en las sienes. (A) bosquejo del cráneo para comparar con la cabeza esquimal en la Fig. 5. (B) Cráneo europeo moderno.
Supongamos, para seguir el argumento, que el hombre antiguo se vio posteriormente obligado a comer alimento duro, después de que la familia inicial se hubiera multiplicado y dispersado; y que este alimento careciese de factores que endureciesen el cráneo en su período de desarrollo formativo; en tal caso el fortalecimiento de los músculos de masticación y cervicales iría de conjunto con la construcción de una superestructura ósea para proporcionar el anclaje necesario en forma de crestas así como de arcos en las zonas frontal, posterior y superior del cráneo, pero el cráneo mismo se mantendría lo suficientemente plástico de modo que sufriría una distorsión considerable. La «quilla» tan evidente en el caso del gorila tendía naturalmente a aparecer en el hombre antiguo debido a que los músculos tiraban de los lados del cráneo hacia el interior, bajo el incremento de la tensión (véase Fig. 8). Cuando la mandíbula se empleaba para romper huesos, etc., el punto principal de fuerza se encontraba generalmente en el mentón, porque la acción de cierre entre los dientes sería normalmente unilateral. Esto de nuevo llevaba a un cierto grado de engrosamiento compensatorio. Pero a diferencia de los simios, el hombre es un ser hablante y hace mucho más uso de la lengua. Hay razones para creer que el refuerzo del mentón humano toma la forma de un arco óseo hacia el exterior en lugar de hacia el interior, debido a esto, y esto le da la prominencia característica de la mandíbula humana. Los simios y otros antropoides, en cambio, reciben el refuerzo en forma de un resalte que se dirige al interior, y esto se conoce como la plataforma símica. En algunos fósiles de hombres antiguos hay alguna evidencia de una plataforma símica, y es de suponer que se trata de un refuerzo adicional a lo que es lo normal para el mentón humano, como compensación para la carga adicional impuesta sobre la estructura en este punto. El tirar de la carne en ausencia de una «cubertería» satisfactoria, o quizá debido simplemente a malas formas a la mesa, contribuyó muy posiblemente al prognatismo alveolar que se encuentra con frecuencia en estos restos antiguos. El creciente desarrollo muscular que surgió bajo el arco zigomático forzó naturalmente este último hacia fuera y exigió una forma más fuerte. Por ello es muy probable que el funcionamiento del mecanismo de la mandíbula determine si el cráneo quedará deprimido o no. Las formas fósiles humanas muestran claramente así que toda la serie ha quedado afectada en un elevado grado por las mismas fuerzas depresivas y compresivas. Así, si el hombre primitivo hubiera quedado totalmente privado de cultura, parece bien cierto que sus restos fósiles hubieran exhibido una cualidad extremadamente primitiva, que se podría interpretar erróneamente con mucha facilidad como prueba de un reciente origen a partir de alguna raza de antropoides. Pero en realidad podría suceder que hubiera sujetos que degenerasen en cualquier período de la historia y que dejasen detrás de ellos un cementerio lleno de los restos fósiles más engañosos. Humphrey Johnson observaba en relación con esto:[65] «Parece
probable que en tiempos muy tempranos la forma humana
poseyera un elevado grado de plasticidad que ha
perdido desde entonces, y que ocasionalmente surgieran
exageraciones de ciertos rasgos raciales,
probablemente ocasionados por un medio ambiente
desfavorable. En la rama Pequín-Java de la familia
humana se ha dado la exageración de los rasgos
simiescos en grado elevado; más adelante tuvo lugar,
por lo que parece, aunque no de forma tan pronunciada,
en el Hombre de Neanderthal, y ha vuelto a ocurrir en
un grado menos acusado con los aborígenes de
Australia.
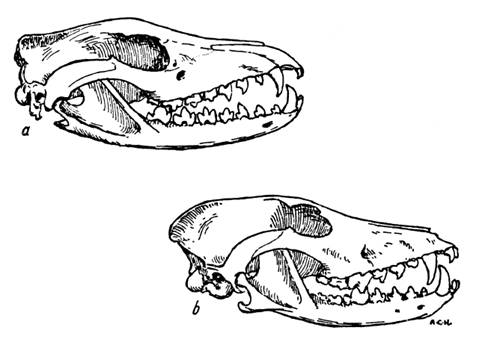
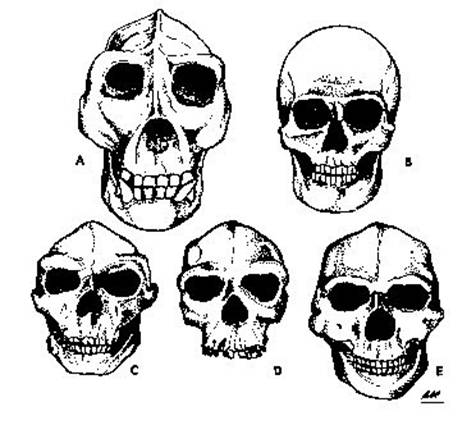
Fig. 7. (A) Gorila, con una marcada cresta y un amplio arco zigomático. (B) El hombre moderno, con una elevada bóveda y la dimensión mayor en las sienes. (C) Pithecanthropus. (D) Hombre de Rodhesia. (E) Sinanthropus.
Fig. 8. Cráneo de gorila hembra (izquierda), Pithecanthropus (centro), y un papú nativo moderno (derecha), vistos desde arriba. Se observan la marcada formación de la arcada superciliar y de la constricción postorbital. Estas destacables diferencias pueden atribuirse casi con toda certeza al desarrollo de potentes músculos para masticar y morder. Si las anteriores interpretaciones son correctas, sigue de ello que un retorno a las condiciones de dieta y vida que caracterizaban al hombre prehistórico irían seguidas de una tendencia a derivar también a su tipo físico. Esta semejanza con los simios que a veces presenta el hombre fósil no tendría nada que ver en absoluto con una descendencia filogenética. En un grupo determinado, el macho adulto tendría más proclividad que el niño (y probablemente más que la hembra) a asemejarse a los simios, pero no se podría sostener en base a esto que el adulto estaba relacionado más de cerca con el ancestro simio que el niño o la hembra. Franz Boas observó lo siguiente hace muchos años:[66] «Si introducimos a dos individuos orgánicamente diferentes en el mismo ambiente, pueden, por ello, volverse parecidos en sus respuestas funcionales, y podemos adquirir la impresión de una semejanza funcional de distintas formas anatómicas que se debe al medio, no a la herencia.» Ya he dicho antes que no tenemos una evidencia inequívoca de que formas simiescas evolucionasen realmente a formas más humanoides, pero sí que tenemos evidencia acerca del hombre volviéndose más simiesco en su estructura craneana. Por cierto que LeGros Clark admitió que este punto preciso se ha propuesto con toda seriedad y con cierta razón:[67] «El Profesor Wood Jones ha observado que hay unos ciertos rasgos anatómicos que facilitan creer que los simios han descendido del hombre en lugar de que el hombre haya descendido de un simio extinto.» Pero él creía que la evolución nunca invierte sus pasos. Por ello, arguyó que nunca ocurriría un paso retrógrado de esta naturaleza. Quizá, pero en forma estructural y por ello, ante una mirada no crítica, el hombre puede «retroceder» parte del camino, aunque el verbo «degenerar» sería mucho más apropiado. El hombre ha sufrido esto, incluso en época relativamente reciente, como señaló Robert Chambers al recoger la historia de ciertos pobres campesinos irlandeses alrededor de 1600 que fueron desposeídos de sus hogares bajo unas circunstancias particularmente duras:[68] «Se
conoce que el estilo de vida tiene un poderoso efecto
en la modificación de la figura humana en el decurso
de generaciones, y esto incluso en su estructura ósea.
Hace unos 200 años, a causa de una política
implacable, un grupo humano fue expulsado de los
condados de Antrim y Down en Irlanda hacia la costa;
desde entonces han estado asentados allí, pero en unas
circunstancias de miseria insólita. Este no es un caso limitado a un solo individuo. Aquí tenemos un caso en el que la apariencia «primitiva» de todo un grupo humano resulta enteramente de factores históricos. Sin duda alguna estaban tan alejados de los simios, por lo que respecta a descendencia, como nosotros ahora, y potencialmente eran tan susceptibles de recibir educación y tan inteligentes como sus coetáneos. Si a esto añadimos casos de individuos aislados que han quedado «proscritos» o perdidos para la sociedad y que sin embargo de alguna manera han sobrevivido hasta la ancianidad, nos encontramos con los ingredientes del «hombre fósil». Por cuanto, debido a la naturaleza del caso, el hombre primitivo comenzó a multiplicarse en un lugar, es de suponer que fuese esparciéndose bajo una cierta presión al irse multiplicando la población. Desde el centro, donde una gran cantidad de población alentaría el desarrollo de una civilización más refinada, irían separándose grupos pequeños como pioneros buscando más espacio y mayor libertad. Los primeros grupos migratorios pequeños hubieran tendido a apartarse más y más del centro hacia la periferia al ir aumentando la presión detrás de ellos, y con ello irían abandonando los refinamientos de la civilización con que empezaron y retrocederían culturalmente, como siempre sucede con los pioneros. Cuanto más elevado su estado inicial, tanto más descenderían en el final. Con el tiempo, las dificultades los llevarían a una mayor disminución de sus recursos, bien porque el grupo fuese demasiado pequeño, o el medio ambiente demasiado duro. Unos pocos supervivientes solitarios podrían finalmente perecer en un terrible aislamiento en el borde fronterizo de las oleadas de emigración, al imposibilitar la ancianidad su continuidad. Estos, en la periferia, pueden ser nuestros «hombres fósiles», descendientes de seres humanos civilizados, no sus antecesores. Arthur Koestler observó con razón[69] que la longevidad bien podría degradar a estos individuos de forma todavía más marcada, hasta que parecieran muy simiescos. La enfermedad puede también jugar un papel. Como sabemos, al primer representante de los Neanderthales se le reconstruyó como un bruto portador de un garrote, encorvado y andando de forma simiesca. En la actualidad se sabe que este primer espécimen era en realidad un individuo enfermo.[70] Se conoce que los hallazgos posteriores de Neanderthales mostraban una postura totalmente erguida.[71] Y los hay que creen que el Sr. Neanderthal podría pasearse por la calle totalmente inadvertido si solo estuviera correctamente vestido y con un corte de cabello apropiado (véase Fig. 9).
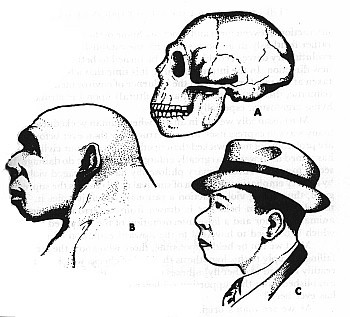
Fig. 9. Este cráneo de Neanderthal (A) de La Chapelle-aux-Saints fue a su tiempo reconstruido (B) para el Museo Field de Historia Natural, Chicago, para mostrar cuál era la apariencia de nuestro primitivo antecesor. Fue reconstruido (C) por J. H. McGregor para mostrar cuán «moderno» puede haber sido realmente en su apariencia.
Respecto a los efectos de las enfermedades, Jesse Williams observaba, en su libro de texto de anatomía y fisiología:[72] «Los tipos degenerados exhiben marcas características que se conocen como estigmas de degeneración. Entre los estigmas comunes están: (1) una frente recesiva, que indica un desarrollo incompleto de los lóbulos frontales del cerebro; (2) prognatismo, una prominencia de los maxilares; (3) el oído canino; (4) arcos superciliares prominentes; (5) pezones situados demasiado arriba y pezones supernumerarios.» Las perturbaciones glandulares pueden tener también unos profundos efectos sobre la anatomía. Keith[73] atribuyó a una pituitaria hiperactiva una tendencia a presentar unos fuertes arcos superciliares, a la vez que una pituitaria subdesarrollada puede explicar una cierta condición plana del rostro de muchos grupos europeos.[74] La conducta cultural puede estar en el origen de cambios marcados en la estructura del cráneo y de la mandíbula. El acto de arrancar carne del hueso en ausencia de cuchillos no solo tiende a fortalecer los músculos maseteros y a alargar algo el arco zigomático como consecuencia directa, sino también a producir una proyección hacia adelante de los dientes frontales, y estos rasgos dan una forma más simiesca a la cara. Por otra parte, tenemos muy poca información precisa acerca de hasta qué punto pueda quedar modificada la estructura ósea en ausencia de una dieta endurecedora de los huesos durante la infancia. Hay mucho espacio para reexaminar toda la cuestión de la interacción entre forma y función. Algo del tiempo perdido de forma totalmente infructuosa en la obsesión por establecer líneas de linajes evolutivos podría quizá dedicarse con mayor provecho en una nueva dirección. En interés de la verdad, es hora ya de que se plantee de una forma nueva toda la cuestión de la influencia de los hechos medioambientales (climáticos, dietéticos, barométricos y culturales) sobre la estructura de los organismos vivientes, especialmente en el caso del hombre. Es totalmente cierto que vivimos en una época en la que la maldad humana encuentra muchas maneras de expresarse de forma más destructiva que nunca en el pasado. Probablemente no es que seamos más malvados que en otras eras, pero nuestra civilización nos ha armado de una manera que multiplica enormemente nuestra capacidad para hacer daño. Me parece que la filosofía evolucionista ha alentado la violencia con su propio énfasis en la idea de la supervivencia a toda costa como el bien supremo, lo que ha dado a la acción violenta un fundamento que la justifica. Este modelo de conducta humana está extraído de una perspectiva errónea de la conducta animal, y de una falsa interpretación de la supuesta dinámica que se cree que ha conducido a la evolución ascendente de toda la vida. ¿No se nos debe considerar responsables, a los que conocemos los hechos, por dejar de aplicar a esta hipótesis los ensayos que aplicamos con todo rigor a otras hipótesis que pueden quedar mejor establecidas en base a la evidencia que las sustenta de lo que jamás lo ha sido la teoría evolucionista? Según consideramos el origen del hombre, así contemplamos su destino. La teoría evolucionista no solo es pésima ciencia, me parece, sino una filosofía todavía más peligrosa. [1] Encyclopedia Britannica, edición de 1964, vol.15, artículo «Morphology», p. 819. [2] Simpson, G. G, This View of Life, Harcourt, Brace & World, Nueva York, 1964. [3] Gerard, Ralph, «The Biological Basis of Imagination», Scientific Monthly, junio de 1947, p. 499. [4] Rudolph Flesch escribió: «Para el lego, lo más importante que debe saber acerca de la ciencia es que no es una búsqueda en pos de la verdad, sino del error. El científico vive en un mundo en el que la verdad (íntegra) es inalcanzable, pero donde siempre es posible encontrar errores en lo largamente aceptado o en lo evidente. ... Los pretendidos libros científicos que se supone que dan respuestas definitivas nunca son científicos. La ciencia es para siempre autocorrectiva y cambiante; ¡lo que se da como verdad absoluta no puede ser ciencia! Citado por Hillier Kreighbaum en Scientific Monthly, abril de 1952, p. 240, de Flesch, The Art of Clear Thinking, Harper, Nueva York, 1951. [5] Evans-Pritchard, E. E., escribió: «toda la historia de la erudición, tanto en las ciencias naturales como en las humanidades, nos dice que la mera recolección de lo que se llama hechos, sin un hilo conductor teórico en la observación y selección, tiene poco valor». Social Anthropology, Cohen and West, Londres, 1951, p. 64. [6] Cf. Washburn, S. L., «The Strategy of Physical Anthropology», en Anthropology Today, dirigido por A. L. Kroeber, University of Chicago Press, 1953, p. 718: «Durante años ha ido aumentando la conciencia de que los hechos por sí mismos no resuelven los problemas, y que incluso la recolección de los “hechos” está dirigida por un complejo conjunto de presuposiciones implícitas». [7] Kropotkin, Prince Petr, Mutual Aid: A Factor in Evolution, Extending Horizon Books, Boston, reimpresión, 1955, xix y 362 pp. [8] Berg, Leo, Nomogenesis: or Evolution Determined by Law (edición rusa original 1922, titulada Nomogenez ili na osnove zakonomernostei), traducción de J. N. Rostovtsov, Massachusetts Institute of Technology, reimpresión, 1969. [9] Manton, I., Problems of Cytology and Evolution in the Pteridophyta, Cambridge University Press, 1950: citado por I. Knoblock, Journal of the American Scientific Affiliation, vol. 5, nº 3, 1953, p. 14. [10] Darwin, Charles, The Origin of Species, 6ª edición, 1872, p. 74. [11] Darwin, Charles, Life and Lectures, 13 de octubre de 1876, iii, p. 159. [12] Gray, Sir James, en una reseña de Evolution in Action, de Huxley, bajo el encabezamiento «The Case for Natural Selection», en Nature, 6 de febrero de 1954, p. 227; y 7 de agosto de 1954, p. 279. [13] De Beer, Sir Gavin, «Embryology and Evolution», en Evolution, dirigido por de Beer, Oxford University Press, 1938, pp. 58, 61. [14] Ibid., p.62. [15] Wallis, Wilson D., «The Structure of Prehistoric Man», en The Making of Man, Modern Library, Random House, New York, 1931, p. 75. [16] Calverton, V. F., en The Making of Man, Modern Library, Random House, New York, 1931, p. 2. [17] Kroeber, A. L., «Evolutionary History and Culture», en Evolution After Darwin, vol.2. University of Chicago Press, 1960, p. 1. [18] Clark, Sir Wilfrid LeGros, Early Forerunners of Man: citado por Rendle Short en Transactions of the Victoria Institute, vol. 66, 1935, p. 255. [19] Clark, Sir Wilfrid LeGros, «The Study of Man's Descent», en A Century of Darwin, dirigido por S. A. Barnett, Heineman, Londres. 1958, p. 182. Hay traducción castellana, Un siglo después de Darwin, Alianza Editorial, Madrid, 1966 – 1971. [20] Weidenreich, Franz, «The Skull of Sinanthropus pekinensis: A Comparative Study on a Primitive Hominid Skull», Paleontologica Sinica, N.S.D., no.10, serie completa nº 127, 1943, p. 1. [21] Zuckerman, Sir Solly, «An Ape or The Ape?» Journal of the Royal Anthropological Institute, vol. 81, 1951, p. 57. [22] Hardy, Sir Alister, The Living Stream, Collins, Londres, 1965, especialmente el capítulo 7, pp. 180ss., «Habit in Relation to Structure». [23] Jones, F. Wood, Trends of Life, Arnold, Londres, 1953, pp. 74, 75. [24] Berg, Leo, Nomogenesis: Or Evolution Determined by Law, p. 174 de la edición inglesa, Constable, Londres, 1926. [25] Berg, Leo, Nomogenesis: Or Evolution Determined by Law, p. 169 de la edición del Instituto de Tecnología de Massachusetts, 1969. [26] Ibid., p. 225. [27] Simpson, G. G., The Meaning of Evolution, Yale University Press, 1952, pp. 183, 184. [28] Shute, Evan, Flaws in the Theory of Evolution, publicación privada, London, Ontario, 1961. [29] Romer, A. S., en Genetics, Paleontology and Evolution, dirigido por Jepson, Mayr, y Simpson, Princeton Univerity Press, 1949, pp. 103ss. [30] Boule, Marcellin, y Henri V. Vallois, Fossil Man, traducido por M. Bullock, Dryden Press, Nueva York, 1957, p. 573, pie de página. [31] Gates, Ruggles R., Human Ancestry from a Genetical Point of View, Harvard University Press, 1948, p. 3. [32] Romer, A. S., en Genetics, Paleontology and Evolution, dirigido por Jepson, Mayr y Simpson, Princeton Univerity Press, 1949, p. 115. [33] Simpson, G. G., Biology and Man, Harcourt, Brace & World, Nueva York, 1969, p. 38. [34] Friedman, Herbert, «Ecological Counterparts in Birds», Scientific Monthly, vol. 63, no. 5, 1946, p. 395, 398. [35] Berg, Leo, Nomogenesis: Or Evolution Determined by Law, edición original rusa de 1922 traducida por J. N. Rostovtov, Massachusetts Institute of Technology Press, reimpresión de 1969, pp. 214, 215. [36] Ibid., p. 224. [37] Manoilov, W. W., en The Medical Gazette, vol.15, 1923: citado por Leo Berg. [38] Berg, Leo, Nomogenesis: Or Evolution Determined by Law, edición original rusa de 1922 traducida por J. N. Rostovtov, Massachusetts Institute of Technology Press, reimpresión de 1969, p. 225. [39] Prosser, C. L., «The Origin after a Century: Prospects for the Future», American Scientist, vol. 47, 1959, p. 539. [40] Berg, Leo, Nomogenesis, etc. p. 223. [41] Ibid., p.221. [42] Short, Rendle A., «Some Recent Literature Concerning the Origin of Man», Transactions of the Victoria Institute, vol. 67, 1935, p. 253. [43] Berg, Leo, Nomogenesis, etc., p. 221. [44] Zawarzin, A. A., Estudios acerca de la histología del sistema nervioso sensorial y de los ganglios ópticos de los insectos (en ruso), San Petersburgo, 1913, vi y 192 pp. [45] Berg, Leo, Nomogenesis, etc., p. 219. [46] Short, Rendle A., «Some Recent Literature Concerning the Origin of Man», Transactions of the Victoria Institute, vol. 67, 1935, p. 253. [47] Jones, F. Wood, Trends of Life, Arnold, Londres, 1953, p. 80. [48] Street, Philip, Animal Weapons, MacGibbon & Kee, Londres, 1971, p. 37. [49] Lack, David, Evolutionary Theory and Christian Belief, Methuen, Londres, 1957, p. 65. [50] Hardy, Sir Alister, The Living Stream, Collins, Londres, 1965, p. 202. [51] Ibid., p. 201. [52] Ibid., p. 200. [53] Simpson, G. G., The Meaning of Life, Yale University Press, 1952, p. 182. [54] Jones, F. Wood, Trends of Life, Arnold, Londres, 1953, p. 71. [55] Berg, Leo, Nomogenesis, etc., reimpresión 1969, p. 241. [56] Ibid., p. 280. [57] Ibid., pp. 281, 282. [58] Ibid., p. 247. [59] Portmann, A., «Das Ursprungsproblem», Eranos-Jahrbuch, 1947, p.19: «Una misma pieza de evidencia adoptará aspectos totalmente diferentes según el ángulo —paleontológico o histórico— desde el que la contemplemos. La veremos bien como un eslabón en una de las muchas series evolucionistas que el paleontólogo intenta establecer, o como algo vinculado con una acción histórica remota. ... Permítaseme decir claramente que, por lo que a mí atañe, los restos del hombre primitivo que conocemos deberían ser juzgados históricamente.» [60] Wallis, Wilson D., «The Structure of Prehistoric Man» en The Making of Man, dirigido por V. F. Calverton, Modern Library, Random House, Nueva York, 1931, pp. 69ss. [61] Ackerknecht, Erwin, Ciba Symposia, vol. 10, nº 1, 1948, p. 912. [62] Howells, William, Mankind So Far, Doubleday, Nueva York, 1945, p. 68. [63] Ibid., p. 131. [64] Hrdlicka, Ales, «Skeletal Remains of Early Man», Smithsonian Instit., Misc. Coll. 83 (1930), p. 367. [65] Johnson, Humphrey, The Bible and the Early History of Mankind, Burns and Oates, Londres, 1947, p. 89. [66] Boas, Franz: citado por Ralph Linton, The Study of Man, Appleton-Century, Nueva York, 1936, p. 26. [67] Clark, Sir Wilfrid LeGros: citado por Rendle Short, en The Transactions of the Royal Victoria Institute, vol. 66, 1935, p. 255. [68] Chambers, Robert, Vestiges of the Natural History of Creation, Churchill, Londres, 1844. [69] Koestler, Arthur, The Ghost in the Machine, Hutchinson, Londres, 1967, p. 167: «La prolongación de la duración absoluta de la vida podría proporcionar la oportunidad para la reaparición de rasgos del primate adulto en los humanos ancianos: Matusalén se convertiría en un simio peludo». [70] El primer espécimen de Neanderthal presentaba evidentes señales de haber padecido osteoartritis crónica, una dolencia que le forzaba a adoptar una postura encorvada. Véase C. S. Coon, The Story of Man, Knopf, Nueva York, 1962, p. 40. Coon dijo que aquellos «huesos estaban podridos de artritis». Véase también la comunicación de A. J. E. Cave ante el 15 Congreso Internacional de Zoología en Londres, reseñado en Discovery, noviembre de 1958, p. 469. [71] Acerca de la postura erguida normal de los Neanderthales, Alberto Carl Blanc y Sergio Sergi, en Science (vol. 90, suplemento, 1939, p. 13). Estos autores comunicaron el hallazgo de dos cráneos adicionales, en los que la base de uno de ellos estaba «bien conservada, lo que hizo posible al Profesor Sergi establecer por primera vez que el Hombre de Neanderthal andaba erguido y no con la postura simiesca con la cabeza proyectada hacia adelante como antes se creía. El plano horizontal de la abertura en el cráneo muestra que los huesos del cuello se ajustaban perpendicularmente en la abertura, lo que hacia que la postura fuese erguida como en el hombre actual.» [72] Williams, Jesse, Textbook of Anatomy and Physiology, 5ª ed., Saunders, Philadelphia, 1935, pie de pág. 49. [73] Citado por Sir John A. Thompson, en The Outline of Science, Vol. 4, Putnam, N.Y., 1922, p. 1097. [74] Keith, Sir Arthur, «Evolution of Human Races in the Light of Hormone Theory», Bulletin of the Johns Hopkins Hospital, 1922. Título: Convergencia, y el
Origen del Hombre Autor: Arthur C.
Custance, Ph. D. Copyright © 2005 Santiago Escuain para la traducción. Se reservan todos los derechos.
© Copyright 2005, SEDIN
- todos los derechos reservados. SEDIN-Servicio
Evangélico |
Vuelta
al Índice de EL PÓRTICO 



|





||| General English Index ||| Coordinadora Creacionista ||| Museo de Máquinas Moleculares ||| ||| Libros recomendados ||| orígenes ||| vida cristiana ||| bibliografía general ||| ||| Temas de actualidad ||| Documentos en PDF (clasificados por temas) ||| |