Robert
Deyes[*]
El
ARN catalítico — un catalizador indigno de una discusión
seria acerca del origen de la vida
Durante las últimas
décadas, la búsqueda de la vida extraterrestre ha sido un
punto focal de la
exploración del espacio. Mientras la idea de seres
extraterrestres en busca de
nuevos planetas habitables sigue generando polémica en los
círculos científicos
y religiosos, más recientemente, en la conferencia de
astrobiología de la
Pontificia Academia de las Ciencias (1), una serie de investigaciones
contemporáneas se ha centrado en los organismos unicelulares
primitivos (2).
Astrobiólogos como Richard Hoover afirman confiados que la vida
primitiva
existe fuera de nuestra propia tierra (3-4). Desde que la nave espacial
de la
NASA Galileo pasó en 1998 cerca de Europa, la luna de
Júpiter, muchos
consideran más que probable la existencia de bacterias en esta
helada luna
(5-6).
La noción de que la
vida podría desarrollarse así sin más, allí
donde se encuentran las condiciones
ambientales apropiadas, exige desde luego un gran salto de fe. Es una
noción
que pasa por alto una multitud de factores críticos, sobre todo
el origen de
alguna clase de material genético codificante. Stephen Mojzsis,
de la
Universidad de Colorado, presentaba la analogía de que el hecho
de que haya un escenario
en un teatro no significa que los actores están presentes y
listos para
desempeñar sus papeles respectivos (7). ¿Qué
procesos habrían tenido que darse
para llevar a una mezcla de compuestos químicos al nivel
requerido de
funcionalidad mínima sobre la que pudiera empezar a actuar la
selección natural
darwinista?
El paleontólogo Niles
Eldredge capturó la relevancia de esta pregunta en su discurso
sobre los ritmos
evolutivos, con su observación de que «hay una enorme
diferencia entre una
colección de moléculas orgánicas incapaces de
captar la energía necesaria para
catalizar su propia replicación, y un sistema organizado que
posee exactamente
esta capacidad» (8). Destacados pensadores, como el
biofísico Paul Davies,
sostienen la idea de que la vida surgió precisamente porque
consiguió
«liberarse de los grilletes de las leyes físicas y del
mundo determinista,
algorítmico» (9). Davies concluye que una aleatoriedad
inicial es la única
manera por la cual podríamos haber llegado a la complejidad
biológica especificada
que encontramos en la vida coetánea (9). No obstante,
¿como hubieran podido
conseguir alcanzar tales actividades unos polímeros moleculares
carentes de
cualquier actividad funcional, solamente por medio de acontecimientos
aleatorios?
En los últimos veinte
años han surgido varias pistas interesantes en el ámbito
del ARN catalítico.
Experimentos realizados a finales de los años 80 y 90 revelaron
que ciertos
tipos de ARN tenían actividades catalíticas
intrínsecas (10). Bioquímicos
renombrados en el estudio del ARN como Tom Cech, Dan Hershlag, Luc
Jaeger y
Anne Marie Pyle proporcionaron detalles clave sobre los procesos que
llevan al
ARN a plegarse en formas catalíticamente activas (10-12). Con la
demostración
de sus capacidades enzimáticas y de soporte de
información, el ARN surgió como
un candidato favorito en la búsqueda de la molécula que
pudiera haber disparado
el tiro de salida del comienzo de la vida (13). El mensaje promulgado
por los
partidarios del «mundo del ARN» era que, mediante una
selección natural
darwinista, las mutaciones pudieron haber producido actividades
catalíticas que
se fueron mejorando a través de generaciones sucesivas de
replicación (13).
Probablemente para
disgusto de los proponentes de la famosa hipótesis sobre el
«mundo de ARN», el
químico de Duke University David Deamer, junto a otros,
desacreditó
convincentemente dicha hipótesis, fundándose en que los
procesos necesarios
para la formación de biopolímeros habrían sido
sumamente ineficaces en una
tierra sin vida. Sus conclusiones resultaron contundentes:
«Está
claro que sería difícil llegar directamente a un mundo de
ARN (o
también a su precursor molecular) a partir de compuestos
orgánicos disueltos en
un océano global (Joyce, 1991). Incluso si hubiese la
posibilidad de generar
nucleótidos químicamente activados capaces de
polimerizarse y de formar ARN en
solución, en ausencia de algún mecanismo que los
concentrase se encontrarían
sumamente diluidos, y no podrían haberse dado ningunas otras
reacciones …
[Dichas] ineficacias inherentes parecen ser inconsecuentes con ir
más allá de
las etapas iniciales de la generación de monómeros y
quizá de unos polímeros
aleatorios.» (13)
Mis propias
investigaciones durante mi estancia en la Universidad de Estrasburgo,
en
Francia, sirvieron para consolidar mi propio escepticismo sobre el
papel del
ARN en el origen de la vida (14). Usando algoritmos que simulan los
pliegues de
ácidos nucleicos, trabajé con otros para diseñar
unas especiales moléculas
catalíticas de ARN llamadas ribozimas, que identificaban de
manera muy
específica secuencias de ARNm dentro de la célula y las
cortaban (véase la
figura abajo; Ref 15). Como pronto descubrí, estas
«tijeras» moleculares no
sólo tenían que cumplir unos estrictos criterios en sus
secuencias para poder discriminar
entre los ARNs adecuados y los no deseados, sino que también
tenían que ser lo
suficientemente cortas para liberarse de sus productos de
reacción y quedar
disponibles para otras rondas de cortes (16). Este último punto
es de suma
importancia si el ribozima ha de exhibir la capacidad de catalizar
repetidas
veces una determinada reacción (17).
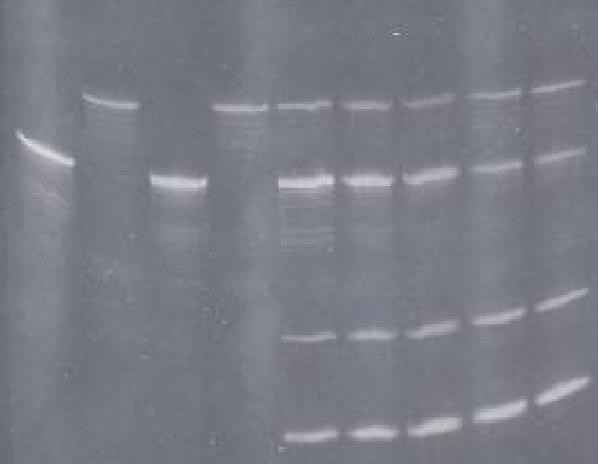
FIGURA: Gel de poliacrilamida 12%
exhibiendo: Carriles 1,3- ARN objetivo; Carriles 2,4-
ARN
ribozima; Carriles 5-7- Curso temporal de la digestión
del ribozima in
vitro (obsérvense los productos de escisión en la
mitad inferior del gel).
Difícilmente se
podría
decir que mi meticuloso diseño de unos ribozimas representa de
manera realista
el proceso de selección darwinista. Si yo no hubiese escogido
mis secuencias
con todo cuidado, no habría obtenido los efectos deseados al
introducir estos
ARN en el interior de las células. Mis propios resultados se
hacían eco de las
conclusiones del biólogo estructuralista Thomas Creighton, que
comentaba que
«la dificultad fundamental con el panorama del mundo del ARN es
que es difícil
explicar cómo dichas moléculas se hubieran podido
sintetizar en una sopa
primordial» (18).
En tanto que diversos
equipos científicos han demostrado diversos tipos de actividad
ribozimática incluyendo
la formación de enlaces péptidos y la ligadura de ARN, el
campo de tales
actividades palidece frente al extenso repertorio de funciones
proteínicas
conocidas (19). Así, ¿en qué medida podemos
considerar suficiente el ARN
catalítico para la formación de componentes que pudieran
posteriormente
ensamblarse para constituir las formas más simples de vida? Por
otra parte, la
realización de tales actividades en el laboratorio sólo
es posible guiando
deliberadamente estas moléculas de ARN hacia puntos funcionales
predeterminados
(19, 20).
Escribiendo en los
años 70, el zoólogo Richard Dawkins compuso el siguiente
«Himno a la causalidad
natural»;
«[La
sopa primordial] debe haber estado poblada por variedades de
moléculas
estables, estables en el sentido de que o bien las moléculas
naturales duraban
mucho tiempo, o bien de que se replicaban rápidamente, o de que
se replicaban
con precisión. Las tendencias evolutivas hacia estas tres clases
de estabilidad
ocurrieron de la siguiente manera: si hubiésemos tomado muestras
de esta sopa
en dos momentos diferentes, la muestra posterior habría
contenido una fracción
más elevada de variedades moleculares con alta
longevidad/fecundidad/fidelidad
de copia. Esto es esencialmente lo que quiere decir un biólogo
cuando está hablando
de seres vivos, y el mecanismo es el mismo —la selección
natural.» (21)
Han transcurrido
treinta años desde entonces, y sigue sin haber carne en los
huesos de los
peregrinos ensueños de Dawkins. Desde el punto de vista del
mundo de ARN, sigue
sin convencerme en absoluto.
Literatura
Citada
1. Tom Chivers
(2009) The Vatican Joins The Search For Alien Life, Véase http://www.telegraph.co.uk/science/space/6536400/The-Vatican-joins-the-search-for-alien-life.html
2. David Malin
(2004) Heaven and Earth: Unseen by the Naked Eye, Phaidon
Press, Reino Unido 2004, p. 284
3. Kate Tobin
(2009) Extremophile Hunter: The search is on for
extremophiles that may provide insights about life elsewhere in the
cosmos,
Véase http://www.nsf.gov/news/special_reports/science_nation/extremophile.jsp
4. Jeff Hecht
(2001) Life will find a way, New Scientist, 17 marzo
2001, p. 4
5. Patrick Barry
(2009) A Tale Of Planetary Woe, Véase http://science.nasa.gov/headlines/y2009/06nov_maven.htm?list207640
6. Clues To
Possible Life On Europa May Lie Buried In Antarctic Ice (1998)
Véase http://science.nasa.gov/newhome/headlines/ast05mar98_1.htm
7. Stephen Mojzsis
habló acerca del origen de la vida en un documental de NOVA que
se emitió en la
cadena PBS el 28 de septiembre de 2004, titulado «Origins: How
Life Began».
8. Niles Eldredge
(1987) Life Pulse: Episodes From The Story of The Fossil Record, Facts
On File
Publications, Nueva York, p. 30
9. Paul Davies
(1999) The Fifth Miracle, The Search for the Origin and The
Meaning of Life, Simon & Schuster, New York, pp. 250-257 (Hay
edición en
español, Paul Davies: El quinto milagro, Planeta-De Agostini,
Barcelona 2004.
10. T. R. Cech
and D. Herschlag (1997) Group I Ribozymes: Substrate
Recognition, Catalytic Strategies and Comparative Mechanistic Analysis,
Nucleic
Acids and Molecular Biology, Vol 10 pp. 1-17
11. L. Jaeger,
F. Mitchel, E. Westhof (1997) The
Structure Of Group I Ribozymes, Nucleic Acids and Molecular Biology,
Vol
10 pp. 33-51
12. A.M. Pyle
(1997) Catalytic Reaction Mechanisms and Structural Features
of Group II Intron Ribozymes, Nucleic Acids and Molecular Biology,
Vol
10 pp. 75-107
13. David
Deamer, Jason Dworkin, Scott Sandford, Max Bernstein, Louis
Allamandola (2002) The First Cell Membranes, Astrobiology, Vol
2 pp.
371-381
14. Robert Deyes
(1998) Observaciones inéditas, Trabajo realizado en LPCCNM-UPRES
2308, Faculté
De Pharmacie, Université Louis Pasteur, Illkirch, Francia
15. Michael Zuker
(2003) Mfold web server for nucleic acid folding and hybridization
prediction, Nucleic
Acids Res, Vol 31 pp. 3406-15 (es una versión actualizada de
la que empleé
en mi investigación)
16. Daniel
Herschlag (1991) Implications Of Ribozyme Kinetics For Targeting
The Cleavage Of Specific RNA Molecules In Vivo: More Isn't Always
Better, Proc.
Natl, Acad, Sci. USA, Vol 88 pp.
6921-6925
17. Thomas
Creighton (1993) Proteins, Structure
and Molecular Properties,
W.H. Freeman and Company, Nueva York, p. 387
18. Ibid, p.107
19. Michael P.
Robertson y William G. Scott (2007) The Structural Basis of
Ribozyme-Catalyzed RNA Assembly, Science, Vol. 315 pp. 1549-1553
20. Gordon C.
Mills y Dean Kenyon (1996) The RNA World: A Critique, Origins
& Design 17:1, See http://www.arn.org/docs/odesign/od171/rnaworld171.htm#note4
21. Richard
Dawkins (1989) The Selfish Gene,
2nd Ed, Oxford University Press, Oxford, Reino
Unido, p. 18.
* Robert Deyes es biólogo
molecular, y ha trabajado
en investigación en biología celular y genética
molecular en centros como la
Universidad de Portsmouth, Reino Unido; Universidad de Atenas, Grecia:
el
Instituto Duncan Guthrie de Genética Médica de la
Universidad de Glasgow,
Escocia; el Departamento de Farmacología de la Universidad Lous
Pasteur, en
Illkirch, Francia. Es autor de siete artículos
científicos en diversas
revistas, como Cell Notes y Neural Notes, entre otras. Ha
participado como representante científico en la Conferencia
sobre Secuenciado y
Análisis del Genoma (2002, 2003 y 2004), en el XIX Simposio
sobre
Identificación Humana (2008) y en la Conferencia de la Sociedad
de Bioquímica
de México sobre Transducción de Señales (2009).
Actualmente vive en Wisconsin,
EE. UU.
Para el original en
inglés, puede acceder a Catalytic
RNA: An Unworthy Catalyst For A Serious 'Origins' Discussion