EL HOMBRE FÓSIL
Frank W.
Cousins
2—EL PROBLEMA BIOLÓGICO Y GENÉTICO
El rocío de tu nacimiento procede del vientre de la
mañana. Sal. 110:3.
Se precisa en el acto de unas palabras de explicación acerca
de la razón de la inclusión de un breve capítulo sobre genética en una obra
dedicada al hombre fósil. Es difícil ver cómo se podría ignorar en cualquier
argumento acerca del linaje del hombre, y sin embargo los huesos fosilizados no
ofrecen la más mínima pista al genetista interesado en las relaciones o
interfertilidad de los dueños de dichos huesos durante su vida. La realidad es
que el argumento acerca de la descendencia del hombre o del quasi-hombre en base
de una cantidad de huesos fosilizados es una mera abstracción e hipótesis—sin
recurso a experimentos de reproducción en los que apoyarse. Las relaciones de
descendencia, en este campo de investigación, se basan forzosamente en el arte
de la anatomía comparativa y en biometría, en muchos casos en la craneometría.
Pero nada de esto da indicación alguna en la búsqueda en pos de una genealogía.
Pero si los huesos yertos no sirven de guía a los genetistas, ¿no deberíamos
cerrar la indagación desde el punto de vista de la genética? No del todo, por
cuanto se afirma muy ampliamente que el argumento en favor de la descendencia
del hombre desde un ser inferior en la escala del reino animal está establecido
como cierto. La evolución
del hombre, nos dicen los evolucionistas, es sólo una pequeña parte de una gran
cadena biológica que comienza desde un primitivo protozoo como la ameba. La
prueba de la evolución del hombre, es decir, de su origen mediante graduales
transformaciones a partir de un protozoo (un animal unicelular) puede
proporcionarse de manera irrefutable sólo mostrando que hay un nexo
ininterrumpido que
vincula a los protozoos con el fílum de los metazoos y de los vertebrados. Para
tener una evidencia completa, se tendría que añadir alguna evidencia que apoyase
al menos que un miembro de los vertebrados se haya transformado en un miembro de
los Hominidae. Nunca se ha podido proporcionar evidencia de ello, y ningún
biólogo ha podido, hasta la fecha (1970), mostrar una sola transformación de un
simple protozoo a un simple metazooni de un vertebrado a
otro.
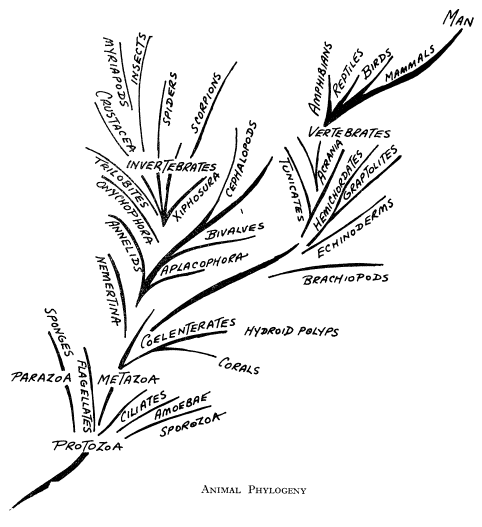
Filogenia
Animal - según de Beer
Este árbol filogenético es típico de la manera en que los
evolucionistas presentan su argumento. Ha sido vuelto a dibujar por el autor
basándose en la obra de de Beer, Atlas of
Evolution [Atlas de Evolución], Nelson, 1964, pág.
155. Al presentarlo, dice de Beer: «Los animales evolucionaron desde las
Protophyta por pérdida de clorofila y adquisición de la nutrición holozoica.
Desde los protozoos, las Parazoos produjeron esponjas y los Metazoos dieron
origen a dos grupos principales que llevaron respectivamente a los invertebrados
superiores y a los vertebrados». No hay evidencia de una cadena evolutiva así.
No hay evidencia al principio de la cadena de que ningún simple protozoo se haya
transformado a un simple metazoo. Este autor no encontró
dificultad alguna en dibujar este árbol filogenético, pero las líneas, las
pendientes de las líneas, el grosor de las líneas, la graciosa curva hacia
arriba de las líneas, no deberían confundirse con una evidencia de verdaderos
vínculos genealógicos.
En tal situación, es a mi juicio una exhibición de descaro
intelectual hablar de «la cadena biológica» o intentar mostrar sólo mediante
dibujos una serie filogenética en la que las líneas «de conexión» o flechas
forman la base entera de los imaginados vínculos. Este es un ejemplo de lo que
el Profesor Thompson denomina, en su crítica del argumento
darwinista, «un escandaloso rebajamiento de las normas científicas».
Así, es una irresponsabilidad de parte de cualquier
antropólogo declarar que la evolución del hombre puede ser demostrada por medio
de un argumento restringido solo a diferencias dentro de la familia Hominidae o
la especie Homo sapiens.
El intento de demostrar la evolución dentro de la familia de los Hominidae es
reprensible por la elemental razón de que la especie es una construcción artificial
—sin validez en la naturaleza, y sin posible recurso a experimentos de
reproducción para su apoyo. Hay poderosas razones para creer, por ejemplo, que
todos los hombres son interfértiles, y que por ello pertenecen a la misma
especie Homo sapiens. El
recurso de los evolucionistas a pequeñas mutaciones dentro de las especies no
puede tener significado para la evolución como tal, por cuanto las pequeñas y
continuas desviaciones desde el tipo original nunca pueden constituir un nuevo
tipo específico —sólo oscilan alrededor del antiguo y nunca se separan de la
especie madre. Lo cierto es que incluso los grandes mutantes (y aun más los
pequeños mutantes) son interfértiles con ella. Por ello, se ve que existe una
incompatibilidad de principio. Y la principal objeción del Profesor Kolliker al
darwinismo en su época fue expresada en estos términos: «No se conocen ningunas
formas de transición entre especies existentes; las variedades, tanto si son
seleccionadas como si son espontáneas nunca llegan tan lejos como para
constituir especies». Desde un punto de vista genético, podemos decir que si nos
retrotraemos en el tiempo, la población humana disminuye más y más hasta que
llegamos a las unidades reproductivas más pequeñas, esto es, un varón y una
hembra humanos. Debido a que el proceso de formación de razas y de especiación
va acompañado de una irreparable pérdida de genes (o de potencial de
recombinación), las especies anteriores deben haber poseído más genes que las
posteriores. Pero esta pérdida no puede seguir de manera indefinida. Por ello,
tenemos que suponer que los genotipos básicos, incluyendo el hombre, aparecieron
en la tierra dotados de un enorme potencial genético (constituyendo así la base
de un fílum) y para este fenómeno es apropiado el término generatio spontanes. «Generación
espontánea», tomado en este sentido, no dice nada acerca del origen absoluto u
original del hombre mismo. No es nada más que un concepto descriptivo que limita
nuestro horizonte teórico de la experiencia (genética).
A menudo se pierde de vista que en tiempos de Darwin el
principal problema de la formación de las especies era si se podría demostrar
que existían diferencias heredables entre las especies. Hace ya mucho tiempo que
este problema quedó resuelto con la obra de Mendel. El problema tiene en la
actualidad un planteamiento muy diferente, y es éste: ¿Se están formando nuevas
especies en la actualidad?
El Profesor H. N. Nilsson señala que sólo dos teorías
intentan proporcionar respuesta al problema y que al mismo tiempo ofrecen una
posibilidad de ensayo. Todas las demás pertenecen al campo de la especulación
filosófica. Las dos teorías son el Lamarckismo y las Mutaciones. Ninguna de las
dos es capaz de «producir» nuevas especies. En los experimentos lamarckianos
acerca de la evolución mediante adaptación, las conclusiones son tan
contundentes que la idea de una adaptación mediante modificaciones heredadas
queda relegada a la oscuridad de la especulación no verificada. Esto no impide a
de Beer,
en una reciente obra de divulgación, mostrar mujeres-jirafa en Birmania y
árboles inclinados por el viento en un capítulo titulado «Acción mecánica que
produce variación fenotípica»; pero de Beer tiene que saber que las
mujeres-jirafa no dan a luz descendencia del tipo jirafa.
El problema de la mutación es complejo, y el lector
interesado debería consultar la fascinante y autorizada obra de
Nilsson.
El problema es enfocado de manera precisa al considerar esta pregunta: ¿Poseen
las mutaciones, tanto las espontáneas como las inducidas, más vitalidad que la
especie madre? Esta es una pregunta fundamental. Las mutaciones pueden ser tan
numerosas como los granos de arena en la playa; puede que se originen de muy
diversas maneras; pero si no pueden competir con su variedad madre en una
población natural, carecen de cualquier valor en el proceso de especiación —y no
puede darse ninguna evolución.
Se puede mostrar, en base de los experimentos de
Gonzales,
Timofeeff-Ressovsky y
Gustafsson, que en ningún caso se puede mantener que ninguno de los mutantes
estudiados tuviera una mayor vitalidad que la especie madre. Por ello mismo, los
mutantes tienen que ser considerados como variedades que sólo pueden existir
bajo condiciones experimentales. Puede que aparezcan en la naturaleza, pero son
inmediatamente eliminados y erradicados. Concluyamos esta breve nota con el
poderoso y provocativo comentario del Profesor Nilsson:
«Una estrecho examen saca a la luz una imposibilidad
empírica inherente en el concepto evolucionista. Si afirmamos que el Homo ha
evolucionado desde la Ameba a lo largo de millones de años, no lo encontramos
imposible, porque las diferencias entre las especies son biológicamente
incomprensibles. Si como eslabones de la cadena escogemos animales específicos
que son algo más similares al hombre, la aceptación no resulta entonces tan
fácil para la mente. La declaración de que el hombre ha sido sucesivamente un
bacalao, un ornitorrinco y un gorila repugna al pensamiento, porque estos
cambios están fuera de nuestra comprensión empírica; pertenecen al mundo de
Alicia en el País de las Maravillas.Si se sugiere que la evolución del hombre ha pasado por etapas de
pez, reptil y simio podemos aceptar la declaración como no imposible, pero en
hecho se trata de una repetición de la misma idea. Se debería tener en mente
siempre el argumento del eminente anatomista K. E. von Baer: “Gradualmente
aprendemos a considerar a las diferentes especies animales como evolucionadas de
otras —y nos olvidamos de que esta metamorfosis es sólo una manera de expresar
los hechos”.»
El estudioso puede aceptar que se le recuerde con algún
mayor detalle la manera en que aparecen las variedades y el gran aumento de
conocimiento que se ha conseguido acerca de esta cuestión desde los tiempos de
Darwin. Por cuanto la observación es que los organismos se reproducen según su
naturaleza, Darwin propuso la doctrina ahora desacreditada de la
pangénesis.
Según esta teoría hay portadores de la herencia, pangenes,
que el torrente sanguíneo lleva a los órganos sexuales y a las células
germinales. Por este medio un cierto órgano, a causa de condiciones externas
cambiadas, adquiere una función más fuerte, de modo que los pangenes aumentan y
se transmiten a las células sexuales en esta forma multiplicada. De ahí, la
progenie obtiene este órgano más poderosamente desarrollado. Estas
características adquiridas, según Darwin, llegan así a ser hereditarias, pero él
siempre había mantenido previamente que el medio no tenía parte alguna en el
proceso evolutivo. Todo esto parece grotesco en la actualidad, en vista del
redescubrimiento por parte de deVries de la obra de Mendel, que él rescató del
olvido. Estos descubrimientos dieron un nuevo aspecto al problema del origen de
las especies, porque nos mostraron que la pauta individual de la especie está
controlada por los genes, que son supremos e inmutables. Así, si dos individuos
tienen unas unidades hereditarias diferentes, estos genes no pueden fundirse en
un nuevo gene en la descendencia. Pero cuando se cruzan dos variedades
diferentes, tiene lugar en la descendencia una recombinación de los genes libres
de modo que se constituyen nuevas variedades. Si los progenitores difieren en 10
rasgos, se pueden formar no menos de 1024 nuevas variedades. El más grande logro
de Mendel reside en haber resuelto el conflicto entre herencia y variación. El
mendelismo nos introduce en un nuevo concepto, por cuanto las formas de
variabilidad que Darwin y otros habían imaginado como emergiendo sucesivamente
unas de otras a lo largo de las eras en una cadena interminable surgen en un
solo momento de uno y el mismo cruce en sus millones como una gran esfera de
variantes. Estas variantes se pueden clasificar según su apariencia en una
serie, pero NO han aparecido en aquella serie. Nosotros clasificamos todo según
la SEMEJANZA, pero la semejanza NO demuestra una semejanza evolutiva, SÓLO una semejanza
constitucional. Nada será
mejor que dejar que la autorizada voz del difunto Profesor Nilsson explique la
historia.
«Así, el mendelismo ha resuelto el problema de la variación
y también el problema de la herencia, pero al mismo tiempo su consecuencias
sobre la teoría de la evolución son similares a las de un peñasco que caiga en
un estrecho riachuelo. Porque el principal resultado del descubrimiento de
Mendel es este: Los genes son
constantes.La variación estácausada por la
recombinación de los genes, no por el cambio de los mismos. Por ello, la
variación queda limitada por las posibilidades combinatorias de los genes. Y
estas quedan limitadas por las posibilidades de cruce. Además, por cuanto los
individuos pertenecientes a diferentes naturalezas de plantas o animales no
pueden siquiera emparejarse, y mucho menos producir progenie, la combinación de
las variaciones queda limitada al interior de la especie. Se forman variantes,
se entrecruzan y vuelven a surgir en una secuencia caleidoscópica
dentrode la especie.
Pero la especie permanece constante como una esfera
de variaciones.Las diversas especies permanecerán
como círculos que no intersectan. Las especies son
constantes. »Con la solución del conflicto entre variación y
herencia, ha surgido un nuevo conflicto, el que existe entre Mendelismo y
evolución. ¿Es grave este conflicto? »Hasta ahora no hemos llegado exactamente a un punto
crítico. Una posibilidad que queda es que se puedan formar nuevas unidades
hereditarias, nuevos genes. ¿Y cómo puede suceder esto? Por mutación. »La aparición de mutantes se ha observado no sólo en
plantas sino también en animales. Y la especie de la que más se habla en la
actualidad, y la más estrechamente estudiada con respecto a su mutabilidad es la
pequeña mosca de la fruta, la Drosophila
melanogaster. »Así, tienen lugar mutaciones, esto es, aquellas formas
de variaciones que no se pueden clasificar en el sistema ordinario de
variaciones mendelianas. Se dice que surgen espontáneamente por un repentino
cambio genético o por un cambio en los elementos portadores de los genes —los
cromosomas— en el núcleo de la célula. Aquí tenemos entonces un factor de una
regeneración totalmente nueva con respecto a las variaciones. Aquí tenemos
nuevas posibilidades de evolución. »Pero —y aquí tenemos realmente otro pero—,
no hay suficiente con que
se formen mutaciones.
Debemos tener la seguridad de que sean causadas por
cambios en los genes, y de que el cambio sea
positivo, en una dirección evolutiva. »Recientes investigaciones han demostrado que en ciertos
casos las mutaciones no se deben a cambios en genes. Y surgen de una manera
totalmente inesperada y totalmente fantástica. Se ha descubierto que ciertas
variedades son, nos sentimos inclinados a decir, una clase de entidades dobles.
También han recibido el nombre de quimeras. Estas variedades se encuentran entre
nuestro pelargonio común. Se las reconoce por sus hojas abigarradas. Si llevamos
a cabo un examen anatómico de una planta con hojas con márgenes blancos,
observaremos que la capa o capas exteriores de células están formadas por
células blancas, pero que debajo hay tejidos verdes. Toda la planta es como una
estructura verde cubierta de un manto blanco. Así, estas variedades han sido
designadas como quimeras periclinales. »Pero sucede que en ciertos casos aparecen ramas con
hojas puramente verdes en estas plantas con márgenes blancos. Se pensaba antes
que había tenido lugar una mutación, en este caso perjudicial. Pero ahora que
conocemos la naturaleza de la planta, el proceso recibe una explicación bien
diferente. »Han causado sensación en años recientes los
experimentos efectuados por el científico americano Muller en el que suscitó
mutaciones en la Drosophila por medio de irradiación con rayos X. Las quimeras (mosaicos) se
conocen también en la Drosophila; lo cierto es que no son cosa infrecuente, y es muy probable que en
este caso la radiación sólo causara perturbaciones en el tejido por medio del
que se diferenciasen constituyentes quiméricos ocultos, de la misma manera que
en el caso anteriormente mencionado. Se descarta que el proceso de mutación sea
de manera general causado por radiaciones naturales, por cuanto no llegan a los
valores cuantitativos precisos para suscitar mutantes experimentales.
En la actualidad es imposible determinar qué
proporción de los mutantes conocidos son causados por verdaderos cambios, y qué
proporción está causada por una diferenciación del tipo de la
quimera. Pero si suponemos que en todo caso algunos
de ellos son mutaciones genéticas espontáneas, siguen permaneciendo algunas
cuestiones que exigen una respuesta, esto es, si van en una dirección positiva y
si tienen un valor selectivo evolutivo. »Por lo que respecta a la primera cuestión, es
destacable el hecho de que la inmensa
mayoría de los mutantes no han adquirido ningún nuevo carácter por
mutación, sino que han perdido uno de los genes de la especie. Son mutantes por pérdida. Para
erigir una evolución con tales mutantes sería necesario suponer que con la
pérdida de genes va asociada una más elevada diferenciación, un desarrollo. La
consecuencia de ello sería que la ameba debería estar dotada con un fondo de
genes enormemente superior al del Homo, y nadie aceptará la consecuencia de tal
punto de vista. Porque entonces los vertebrados serían, en su dotación genética,
sólo amebas muy adelgazadas. »Sin embargo, una proporción muy pequeña de mutantes parecen haber
sido enriquecidos con un gene, parecen ser mutantes
progresivos. Y en el caso de los mismos será
entonces de gran importancia la última prueba: ¿Tienen un valor selectivo que
les capacite no sólo para prevalecer en la lucha por la existencia sino también
para sustituir a la especie progenitora? »Por lo que respecta a la viabilidad, los mutantes están
por lo general mal equipados. No menos de 60 de los 400 mutantes de la
Drosophila han obtenido
el nuevo carácter para perecer. Las plantas deficientes en clorofila y
diabéticas y moscas con alas inútiles o con patas partidas están naturalmente
condenadas a una rápida eliminación. Pero incluso los mutantes menos divergentes
presentan una constitución más débil y una menor longevidad que la especie
progenitora. Esto aparece de manera clara y concluyente en el hecho de que
no se encuentran en la naturaleza mutantes de
Oenothera ni de Drosophila. Naturalmente, aparecen,
y se pueden encontrar casualmente, pero pronto son eliminadas y no forman un
componente del fondo de la naturaleza, y mucho menos puede sustituir la especie
actual. Evidencian ser sencillamente variantes no
viables, y que podemos ver sólo en ciertas circunstancias favorables de cultivo,
pero que en la naturaleza son rápidamente barridas y desaparecen sin dejar
rastro. Y esto es cierto de los mutantes dominantes
lo mismo que de los mutantes por pérdida. Como regla, se puede decir que
cuanto más divergente sea un mutante, menos viable
es. Intentar edificar una evolución en base de
mutantes es más que difícil. Es como lanzarse a un mar tempestuoso en un
cascarón. »Es evidente que las investigaciones de las últimas tres
décadas acerca del problema del origen de las especies no han podido mostrar que
se forme mediante mutaciones un material de variación capaz de competición en la
lucha por la existencia. Además, ha resultado también imposible demostrar una
adaptación progresiva por medio de la transmisión de los caracteres adquiridos
(todos los numerosos experimentos efectuados han dado resultados negativos). Por
ello, somos llevados a la conclusión de que la teoría
de la evolución no ha sido verificada por investigaciones experimentales acerca
del origen de las especies.»
Este comentario, que procede de una autoridad mundial en
paleobiología, fue hecho ya hace treinta años, pero, ¿sabe alguien de ninguna
evidencia que lo refute?
Que cada lector examine su posición intelectual con
sinceridad. Si su estudio acerca del tema es superficial, que examine lo que los
biólogos consideran como la obra más erudita disponible acerca del tema: Me
refiero a la obra del doctor Ernst Mayr, Animal
Species and Evolution, de Belknap Press, de Harvard
University Press (Cambridge, Massachusetts, 1965). Verá allí la deficiencia del
argumento evolucionista, por cuanto el doctor Mayr no trata acerca de los
argumentos del Profesor Nilsson, pero declara (en las págs. 174-181)
que
«No se debe olvidar que las mutaciones son la fuente última
de todala variación
genética que se encuentra en las poblaciones naturales y que es la
únicamateria prima sobre
la que puede trabajar la selección natural.»
En base de mi propio estudio acerca del problema, tengo la
certidumbre de que no se conoce ningún mecanismo satisfactorio para la evolución
y que esta hipótesis se mantiene por razones filosóficas.
Este punto de vista es ampliamente sustentado por la
contribución de la doctora Marjorie Grene a Encounter, Vol. XIII, págs. 48-56,
titulado «The Faith of Darwinism». Doy abajo un extracto procedente de su
análisis de esta posición.
«Ni el origen ni la persistencia de los grandes nuevos
modos de vida —la fotosíntesis, la respiración, el pensamiento—ni todos los
intrincados y coordinados cambios necesarios para soportarlos, se explican o
siquiera se hacen concebibles en el sistema darwinista. Y si uno pasa a leer el
Origenencuentra, por
cierto, que por mucha brillantez que tengan sus hipótesis amontonadas sobre
hipótesis, que a pesar de toda la espléndida simplicidad del “mecanismo”por el
que “explica”tantos y diversos fenómenos, simplemente no trata del origen de
las especies, y mucho menos los grandes órdenes, clases, filas, en absoluto. Su
argumento se desplaza en una dirección totalmente diferente, en dirección de
diminutas adaptaciones especializadas que no conducen a ninguna parte, como no
sea a la extinción. Lo mismo sucede con toda la inmensa, infinita e ingeniosa
montaña de trabajo de los actuales darwinistas: C’est
magnifique, mais ce n’est pas la guerre!Es bien
posible que el color de las polillas o de los caracoles o que la flor en el
tallo del ricino se “expliquen”mediante mutación y selección natural; pero lo
que la teoría neodarwinista deja sin tratar es precisamente la cuestión de cómo
a partir de unos antecesores unicelulares (y desde la misma materia inanimada)
surgieron el ricino, las polillas y los caracoles, y cómo a partir de éstos
surgieron llamas y erizos, leones y simios —y hombres. Con un ingenio infinito
se han elaborado las condiciones microscópicas para estos acontecimientos
macroscópicos; pero no proporciona ningún marco conceptual en términos del cual
se pueda admitir su existencia, y mucho menos una “explicación”de su
descendencia procedentes de formas “inferiores”. En resumen, las reflexiones
acerca de algunos de los problemas de la microevolución pueden bien conducir a
observaciones como la del Profesor Waddington, arropada en los pliegues de su
ingenioso y “ortodoxo”argumento acerca de La Estrategia de los Genes
(The Strategy of the Genes):“... el estudiante carente de
prejuicios podrá recibir la impresión de que el fracaso de la teoría actual en
proporcionar ninguna explicación plausible para tales acontecimientos ha tenido
una parte importante en hacer bajar la balanza en contra de la aceptación de su
existencia real. Parecería de cierto que en este campo ... se pueda dudar de la
idoneidad de la moderna teoría.” »Además, los evolucionistas escépticos de la síntesis
neodarwinista tienen evidencias empíricas para dar apoyo a sus dudas. Porque a
pesar de las pretensiones neodarwinistas, hay dos grandes disciplinas
biológicas, la paleontología y la embriología, que parecen aportar su principal
peso contra el dogma seleccionista.»
|
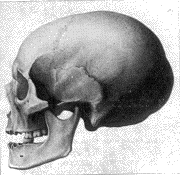
1. Norma
lateralis |
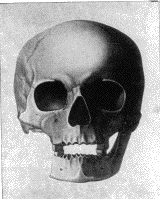
2. Norma
facialis |
1 y 2. Un cráneo mesocéfalo de
Furfooz.
|
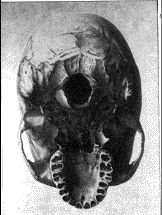
3. Un cráneo dolicocéfalo
Norma basalis. |
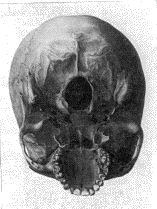
4. Un cráneo braquicéfalo.
Norma basalis.
|
Estas cuatro ilustraciones están tomadas de la magnífica
obra por de Quatrefages y Hamy, CRANIA ETNICA (París, 1882). En la parte
superior (1 y 2) se exhibe un cráneo de varón de la cueva Trou de Frontal en
Furfooz, una aldea a unos 16 kilómetros de Dinant, en las Ardenas. Allí se
excavaron, en 1872, tres cuevas conteniendo restos prehistóricos. Es un cráneo
mesocéfalo producido aparentemente por el cruce de antecesores braquicéfalos y
dolicocéfalos.
En los cráneos de la parte inferior (3, 4) se observa
claramente el foramen magnum. El cráneo (3) es dolicocéfalo y pertenecía a un habitante de Nueva
Guinea. El cráneo (4) es braquicéfalo y perteneció a un habitante de
Córcega.
SEDIN
Servicio Evangélico - Documentación -
Información
Apartat 2002
08200 SABADELL
(Barcelona) ESPAÑA
|
Índice:
Índice de
boletines
Índice
de línea
sobre línea
Página
principal
Índice
general castellano
Libros recomendados
orígenes
vida
cristiana
bibliografía
general
Coordinadora
Creacionista
Museo de
Máquinas Moleculares
Temas de
actualidad
Documentos en
PDF
(clasificados por temas)
Para descargar el
libro en formato
PDF
para Acrobat Reader,
haga clic aquí (1,5 Mb)




|