William E.
James, Ph. D.
Santiago Escuain
El origen
del primer sistema vivo
ALTERNATIVAS
Hay
sólo dos alternativas por lo que respecta al origen de la vida:
a) Designio - ¿Fue la vida creada por medio de actos
inteligentes?
b) Azar - ¿Fue la vida originada por procesos
aleatorios, casuales?
¿EJEMPLOS
PRÁCTICOS?
He
aquí unos ejemplos que surgen de la observación diaria:
a) ¿De las partículas al hombre por medio de
cambios graduales, o de los patines al Rolls Royce por designio?
b) ¿Emergencia de un reloj de pulsera a partir de sus
elementos metálicos por ventajas selectivas durante un largo
período de tiempo; o diseño de un cronómetro
gracias a la entrada de información y de energía
dirigidas a la consecución de un propósito
específico?
Se
debe observar que la célula es un intrincado mecanismo,
y que el origen de la primera célula viviente puede ser
verdaderamente considerado de manera paralela a la del origen de un
reloj. La mera negación de la evidencia del designio no
demuestra en absoluto la inexistencia del designio y fin propuesto para
la vida.
¿POR DÓNDE EMPEZAMOS?
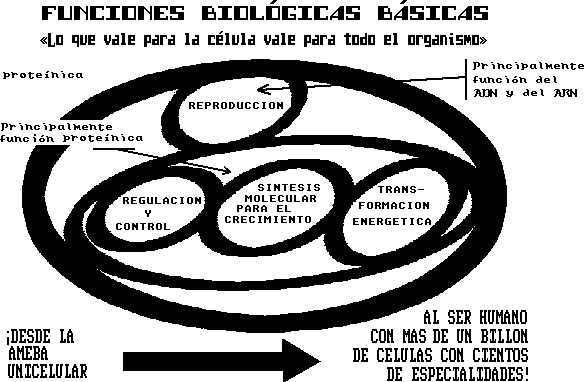
La
unidad viviente básica es la célula viva. La
célula está compuesta de moléculas funcionales,
las cuales:
a) Transforman la energía en formas útiles
para mantener la organización consistente con la vida.
b) Regulan y controlan el medio interior.
c) Controlan el modelo de reproducción celular.
d) Sintetizan las moléculas funcionales.
COMENZANDO POR EL PRINCIPIO
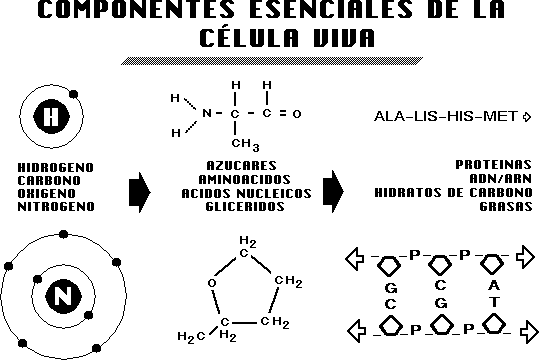
Los
átomos, al combinarse, forman moléculas sencillas; las
moléculas sencillas, al combinarse, forman las moléculas
inmensamente complejas que contiene la célula viviente.
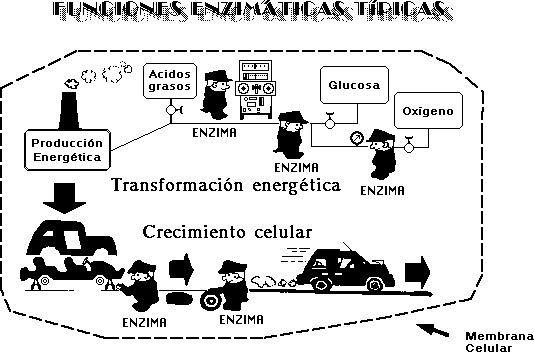
¿CÓMO ES QUE FUNCIONA?
Las
proteínas actúan en cada fase de las ordenadas funciones
de las células.
a) Los enzimas (proteínas) dirigen y controlan la
descomposición de moléculas de alta energía, como
la glucosa, para utilizar esta energía en las operaciones de la
célula.
b) Muchos enzimas pueden ser inhibidos a fin de evitar la
producción de subproductos que serían perjudiciales para
la operación de la célula. Efectúan una
regulación y un control esenciales.
c) Los enzimas tienen actividad en la síntesis de
moléculas sencillas, la cual se emplea para integrarlas en las
estructuras moleculares complejas funcionales de la célula
viviente.
d) Los enzimas suplementan la acción del ADN y del
ARN durante la transcripción de información que tiene
lugar en el proceso de reproducción.
e) Las proteínas tienen la actividad de controlar la
entrada y salida de materiales de la célula.
DE LA «A» A LA «Z»
Para
que las proteínas sean funcionales deben ser estructuradas en la
secuencia correcta. Se acepta generalmente que la variación
permisible en la estructura para que la proteína permanezca
funcional es muy pequeña, si es que puede haber alguna en
absoluto. Se ha determinado que la cadena de aminoácidos debe
estar doblada precisamente de una manera muy específica a fin de
que tenga lugar su funcionalidad. Es la secuencia de aminoácidos
la que determina la manera exacta en que se doblará la
proteína.
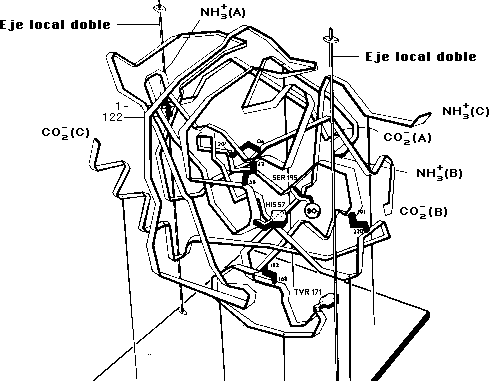
Modelo de la alfa-quimotripsina mono-0-tosilada en la Ser-195

LA DELICIA DEL PROBABILISTA
La
secuencia de aminoácidos le da al matemático oportunidad
de calcular la probabilidad de que el verdadero orden surja al azar.
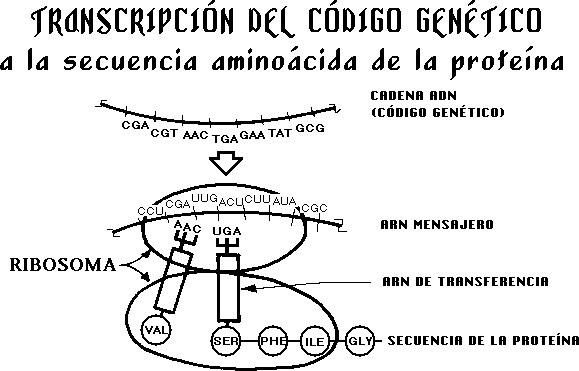
EL MAPA DE CARRETERAS
La
base química del código genético fue determinada
por los premios Nóbel Watson y Crick a comienzos de la
década de los cincuenta. La molécula ADN contiene
ácidos en secuencia, que forman lo que se llama
«codones». Cada «codón» representa una
letra del «alfabeto» genético. Este alfabeto dicta
una especificación completa en forma de código que
contiene la información necesaria para el diseño,
construcción, operación y mantenimiento de toda la
célula. Este código es tan extenso que si se
mecanografiara cada secuencia como una letra del alfabeto, se
necesitaría toda una biblioteca para que contuviera los libros
que se llenarían para describir la célula más
sencilla. Se precisaría de muchos miles de volúmenes para
poder contener todas las letras de la especificación de los
seres humanos. Y con todo, se halla todo el código
genético en cada célula del cuerpo humano (billones de
ellas).
INSTRUCCIONES ESCRITAS
Las
proteínas son «ensambladas» dentro de las
células a partir de aminoácidos, consiguiéndose la
secuencia correcta a partir del código almacenado en la
molécula ADN y, por tanto, el ADN es el responsable de la
secuencia aminoácida de todas las proteínas.
Parece que, según el evolucionismo, la molécula ADN
hubiera debido desarrollarse la primera, antes que hubieran podido
formarse las proteínas. Pero sea que fueran cientos de
proteínas, o una molécula de ADN con millones de codones,
no hay mucha diferencia: la probabilidad de que ello ocurriera es
insignificantemente pequeña.
Carl
R. Woese, en «Progress in Molecular and Subcellular
Biology» (1969) estimaba que para que una célula
bacteriana funcione normalmente, el código ADN debe
transcribirse con un error inferior a una por cada 10.000 a 1.000.000
de letras del código genético.
En
la actualidad se propone generalmente que estas moléculas, tan
complejas, se unieron en el seno de un «caldo primordial»
gracias a los millones y millones de años que transcurrieron.
George Wald, en un artículo titulado «The Origin of
Life» (El Origen de la Vida) en Scientific American de
agosto de 1954, hace la siguiente afirmación:
El tiempo con que tenemos que ver es del orden de dos mil
millones de años. Con tanto tiempo disponible, lo
«imposible» se transforma en posible, lo posible probable,
y lo probable virtualmente cierto. Uno sólo tiene que esperar;
el tiempo por sí solo hace los «milagros».
¿A QUIÉN LE PREOCUPA EL TIEMPO?
Consideremos
el factor tiempo. Los fisicoquímicos están conformes en
que los átomos y las moléculas de gas viajan
rápidamente por el espacio, y que estas partículas chocan
unas con otras, reaccionando para formar moléculas más
complejas. Conociendo el número de moléculas de gas en un
espacio determinado, su diámetro y la velocidad a la que se
desplazan, se puede calcular el número de colisiones por
segundo. Aunque los líquidos no son tan bien conocidos, siguen
estas reglas de una manera sorprendente. Utilizando este enfoque, es
posible calcular el número de colisiones totales de las
moléculas en el mar primordial durante miles de millones de
años. Esto nos daría la cantidad máxima de
combinaciones moleculares que podrían tener lugar durante el
tiempo demandado por el evolucionismo.
¿CALDO O SALSA?
La
Teoría Neodarwinista de la Evolución propone que el
primer organismo viviente tuvo su origen en el mar primordial. Se
supone para ello que este mar contenía una solución
bastante concentrada de aminoácidos y de ácidos
nucleicos, con lo que se formaron las proteínas y el ADN para
formar las primeras «moléculas vivientes».
Recientemente, esta teoría ha sido desafiada por el bien
conocido evolucionista Klaus Dose, en Origin of Life and
Evolutionary Biochemistry, Plenum Press, 1974. Hace él la
siguiente afirmación:
La concentración en equilibrio de estos
dipéptidos para una solución molar de aminoácidos
es así tan sólo ligeramente superior a 10E-5 molar. La
barrera termodinámica es aún mayor para la
formación de un polipéptido o de una molécula de
proteína en solución acuosa. Por lo tanto, parece
evidente que la concentración media de aminoácidos libres
en el océano primordial nunca fue significativamente diferente
de su nivel actual.
El
doctor Dose concluye señalando que los aminoácidos y
proteínas, los ladrillos esenciales de la vida, no
podrían darse en concentraciones mayores que las que observamos
en los océanos actuales. Esto a pesar de las condiciones ideales
que se han postulado para la tierra primitiva. Nadie propondrá
que la concentración actual del océano sea suficiente
para que se origine allí la vida. En nuestros cálculos
hemos empleado la muy generosa concentración de
aminoácidos de 1 molar. Esta concentración es 100.000
veces mayor que la que sugiere Dose.
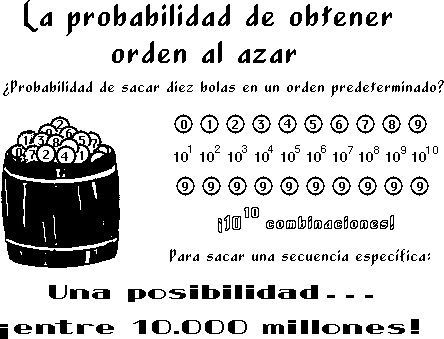
CALCULANDO LAS PROBABILIDADES
La
probabilidad de que apareciera una secuencia precisa de
aminoácido en el mar primordial durante un período de 10
mil millones de años es de una entre 10E420 (un diez seguido de
420 ceros). Casi todos los bioquímicos aceptan que solamente se
puede alterar un pequeño porcentaje de la secuencia de
aminoácidos sin inutilizar la funcionalidad de la
proteína. Citando de nuevo al doctor George Wald, esta vez en un
Simposio convocado por el Instituto Wistar de Anatomía y
Biología:
Habría mucha dificultad en hallar un solo caso en el
que el cambio de un solo aminoácido de su secuencia no cambie
notablemente las propiedades de la proteína.
Si
suponemos que sólo la mitad de los aminoácidos en una
proteína tiene que estar en una secuencia correcta, la
probabilidad sería de una entre 10E185. Si suponemos que
sólo una cuarta parte de los aminoácidos tuviera que
estar en la secuencia correcta, la probabilidad llegaría a ser
de una entre 10E55. Si consideramos que en la célula más
sencilla existen 1.500 proteínas y un millón de genes, el
número se agranda hasta más allá de toda nuestra
capacidad de comprensión. Esencialmente, el exponente
estaría en el orden de los millones, o sea, un diez seguido de
millones de ceros.
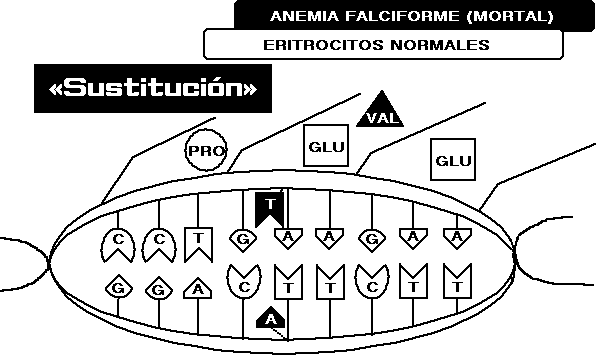
En
realidad, ¿son factibles tantas sustituciones? Contemplemos un
sistema complejo como el organismo humano: Si en la sección de
la doble hélice que gobierna la síntesis de la
proteína hemoglobina se da una mera traslocación
de modo que un par A-T (enfrentados entre
sí a cada lado de la doble espiral) se invierte y pasa a ser
T-A, los codones respectivos pasan a ser CAT y GTA en lugar de CTT y
GAT. Esto da el mensaje de «Valina» en lugar de
«Acido glutámico», y la cadena de hemoglobina
resultante tiene una forma distinta.
Si
no existe una información correcta en el organismo, que
enmascararía este error, esta instrucción es la empleada
para la hemoglobina, que da como resultado unos eritrocitos de
forma de hoz, con lo que el individuo muere debido a la llamada anemia
falciforme. La mera traslocación de dos bases en el
código genético en una larguísima cadena de ADN
lleva a un error de un solo aminoácido en la cadena de
la hemoglobina, y, como resultado lógico, la muerte del que
sufre esta dolencia.
Se
debe observar que las sustituciones de bases en los codones del
ADN no siempre comportan un cambio de aminoácido en la cadena
proteínica. Esto se debe a que varios de los aminoácidos
están codificados por dos y hasta tres grupos
distintos de bases. Esto da lugar a las llamadas mutaciones neutras,
en las que hay un cambio de base que por dar lugar a un codón
que codifica al mismo aminoácido, no hay cambio en el resultado
final, la proteína codificada por el gene respectivo. Este es
uno de los varios mecanismos que ayudan a mantener la estabilidad del
organismo frente a cambios y a accidentes. Hay otros, como el
maravilloso de la reparación enzimática, el cual
detecta en muchos casos los daños sufridos por la cadena de ADN,
y la repara devolviéndola a su estado original.
Lo
dicho sirve como ilustración de lo crítica que es la
disposición de los aminoácidos en las cadenas
proteínicas y que, en todo caso, la concesión de un 10
por ciento de sustitución es extremadamente generosa, por no
hablar un cincuenta o incluso un setenta y cinco por ciento de
sustitución. E incluso admitiendo esta cantidad imposible de
elasticidad de sustitución de un 75 por ciento de
aminoácidos en una cadena de proteína,
¡habría sólo una probabilidad entre 10E55, un
número 100.000 veces mayor que el umbral de imposibilidad
cósmica!
EL MICROORGANISMO DE
MOROWITZ
¿Cuál
sería la probabilidad para el origen de un primer sistema vivo
sumamente simple?
El
organismo mas simple conocido es el Mycoplasma Hominis H39, que
tiene alrededor de 600 clases diferentes de proteínas [Journal
of Molecular Biology, Vol. 23 (1967), pág. 198]. En base de
nuestros conocimientos científicos actuales no hay razón
alguna para postular que jamás haya existido ningún
sistema viviente más pequeño. Sin embargo, seguiremos las
cifras del doctor Morowitz que, trabajando para la NASA,
desarrolló unos cálculos para determinar el límite
teórico inferior para el más simple sistema vivo capaz de
reproducirse. En sus cálculos tuvo en consideración los
mínimos de sistema operativo y el espacio que éste
exigiría, así como las propiedades eléctricas y
los peligros de la agitación térmica. Por sus importantes
estudios llegó a la conclusión de que el más
pequeño sistema teórico posible exigiría 239 o
más moléculas de proteína individuales incluyendo
al menos 124 diferentes especies de proteínas [citado en
Coppedge, J. F.: Evolution: Possible or Impossible?, Zondervan
(Grand Rapids 1973), pág. 72, 110].
El
doctor Coppedge da unas facilidades aún mucho más
generosas a los cálculos del origen de la vida por azar que los
dados hasta aquí. Por ejemplo: (1) Él comienza con toda
la corteza terrestre compuesta enteramente de los isómeros
levógiros de los veinte alfa-aminoácidos, protegidos de
destrucción, (2) asigna a cada intento de formación de
cadena un tiempo de sólo un tercio de billonésima de
segundo (en el seno de un sistema metabólico, como en el Escherichia
coli, se precisa de 5 segundos para una cadena de proteína
de 400 unidades de longitud, a la temperatura óptima de 37oC),
etc. El resultado de sus cálculos es que sólo
habría habido 1 posibilidad entre 10E161 para la
formación de una sola proteína biológicamente
útil. Esto da una idea de lo muy favorables que son sus puntos
de partida para un hipotético origen de la vida al azar, siendo
que en los que hemos dado aquí, también partiendo de
presupuestos enormemente favorables a un origen de la vida por azar, se
ha llegado a la cifra de 1 posibilidad entre 10E420.
Partiendo
de la probabilidad calculada por el doctor Coppedge para una
proteína, ¿cuál sería la probabilidad
de que surgieran las 239 proteínas específicas necesarias
para el más simple microorganismo de Morowitz? Recuérdese
que nos limitamos a las proteínas en sí, y que
dejamos de lado el origen de su ADN y ARN. El cálculo para este
conjunto de 239 proteínas arroja una probabilidad de 1 entre
10E122470. Incluso admitiendo que un billón de secuencias
diferentes pudieran ser funcionales en cada proteína, la
probabilidad resultante para el origen de las proteínas
necesarias para el sistema biológico de Morowitz es de 1 en
10E119775 para toda la edad de la tierra (Coppedge, op. cit.,
pág. 111). Este número nos expresa solamente la
probabilidad de conseguir al azar el conjunto de proteínas,
sino que queda el problema de que se encuentren en el tiempo y en el
espacio, y de su organización y concatenación para
constituir el mencionado sistema vivo.
Desde
otra óptica, el doctor Morowitz llevó a cabo un completo
estudio termodinámico de las células vivientes, empleando
los resultados como base para estimar la probabilidad de la
abiogénesis. La probabilidad termodinámica de la
formación de un organismo como el Escherichia coli sin
dirección, al azar, es una entre 10E34.000.000.000 [Harold
Morowitz, Energy Flow in Biology (Academic Press, N.Y., 1968),
págs. 98-99]. Se ha intentado esquivar este problema (1)
proponiendo fluctuaciones en un sistema que se aparta del equilibrio, o
(2) apelando a un sistema abierto con influjo de energía. Pero
esas propuestas no han quedado sustanciadas en absoluto. Por una parte,
precisamente lo que se está evaluando es la probabilidad de
tales fluctuaciones. Por otra parte, se debe observar que de nada
sirve un mero influjo de energía no dirigida en el
sistema. La síntesis proteínica tiene lugar en el mundo
de lo viviente mediante un complejo dispositivo de conversión
energética, que captura esta energía mediante la
función clorofílica, de enorme complejidad. Esta
energía se transmite a través de la cadena trófica
(de alimentación) y se aplica mediante intrincados procesos
celulares de enorme precisión. El influjo de energía no
dirigida, en ausencia de unos mecanismos determinados por una
información, no conduciría al logro de ningún
orden. Esto podría ilustrarse con la cantidad de energía
necesaria para ordenar las figurillas de porcelana en un
almacén. Nada se ganará soltando un toro bravo dentro del
mismo. Por otra parte, si se da un criterio ordenador a una o varias
personas, éstas aplicarán su energía,
constreñida por unos sistemas de conversión
energética que irán dirigidos por una información.
Así, un sistema abierto es ciertamente una
condición necesaria, pero no suficiente. Por
sí mismo es totalmente irrelevante.
¿CUÁN GRANDE
ES GRANDE?
Para
ilustrar la magnitud de estos números, imaginemos que toda la
tierra estuviera hecha de canicas. El globo terrestre
consistiría de aproximadamente 10E24 canicas. Si todas las
canicas fueran del mismo color excepto una de ellas, la probabilidad de
que un hombre con los ojos vendados la encontrara al primer intento
sería de una entre 10E24. Los estadísticos dicen que esto
está fuera del «umbral de la posibilidad». Y la
mayor parte de nosotros estará de acuerdo.
Si
pasamos del 10E24 a 10E25 parece que sólo hemos aumentado un
poco el número. En realidad, 10E25 es un número diez
veces más grande que 10E24, y en lugar de ser un globo terrestre
compuesto de canicas, tendríamos 10 globos terrestres.
Si llegamos al número 10E28, estamos hablando de diez mil
globos terrestres. ¿Qué diremos de un número como
10E119775?
ÉMIL BOREL Y LOS
DISTINTOS GRADOS DE IMPOSIBILIDAD
En
su libro Las probabilidades y la vida (Ediciones Orbis,
Barcelona 1986), Émil Borel determina las magnitudes de los
diferentes umbrales de imposibilidad para sucesos en diferentes
entornos. En el Capítulo III de la obra (págs. 37-59)
clasifica así los umbrales de imposibilidad: 1) A escala humana;
2) a escala terrestre; 3) a escala cósmica; 4) a escala
supercósmica. En la escala cósmica, esto es, a
nivel de todo el universo existente, determina Borel que «un
fenómeno cuya probabilidad es 10E-50 [es decir, 1 entre 10E50]
nunca se producirá o, por lo menos, nunca será
observado» (op. cit., pág. 42). La escala
supercósmica es hipotética, y simplemente trata de
establecer un límite en un ámbito absolutamente e
inimaginablemente colosal. Acerca de la escala supercósmica,
dice Borel: «Imaginemos, pues, con Boltzmann, un Universo U2 que
abarcara tantos universos U1 análogos al nuestro como
número de átomos posee; luego, un Universo U3 que
encerrara tantos universos U2 como número de átomos posee
U1; después, un Universo U4 que contuviera tantos U3 como
número de átomos posee U1, y así sucesivamente,
repitiendo un millón de veces la misma operación, es
decir, hasta un Universo U[N], con N = 10E6. Este superuniverso
contendría un número de átomos igual a 10 elevado
a la potencia de 110 millones, o sea, que estaría representado
por un número de 110 millones de cifras ...» (op. cit.
pág. 44). Luego Borel hace un razonamiento análogo con el
factor tiempo. Todo esto da un número total de acontecimientos inferior
a 10 elevado a la potencia de 10E9, que se considera como el umbral de
imposibilidad a escala de este hipotético supercosmos.
Hemos
visto que a escala cósmica es imposible todo
acontecimiento con una probabilidad igual o inferior a 1 entre 10E50. Y
en realidad, deberíamos considerar también que a escala
terrestre el umbral de imposibilidad se encuentra en 10E-15, esto es,
que un suceso de probabilidad igual o inferior a 1 entre 10E15 no
sucederá a escala terrestre. La probabilidad del surgimiento de
las proteínas necesarias para el hipotético sistema vivo
simple de Morowitz es de 10E119775, considerando (1) que para llegar a
esta cifra se han hecho muchas concesiones absolutamente irrazonables
al proceso de azar y (2) que con ello sólo tenemos el
conjunto de proteínas, pero no en absoluto que se hayan
encontrado en el tiempo y en el espacio, y que se hayan organizado y
concatenado para constituir el mencionado sistema vivo.
CONSTRUYENDO SOBRE UN
FUNDAMENTO DETERMINADO
¿Por
qué, entonces, se acepta la teoría de la
evolución? Algunas de las razones dadas por aquellos que aceptan
la evolución son puntos de vista filosóficos que excluyen
de entrada lo sobrenatural. El doctor Richard Lewontin, orador en el
«Simposio de Desafíos Matemáticos a la
Interpretación Neodarwinista de la Evolución»,
convocado por el Instituto Wistar de Anatomía y
Biología, afirmó lo siguiente:
Creemos que el error metafísico fundamental que se
está defendiendo aquí es el de decir que si un conjunto
de posibilidades aceptables presenta una magnitud cercana a cero, en
relación con la magnitud total del espacio, la vida nunca
hubiera surgido. Pero el hecho es que la vida surgió, y puede
que haya sido un acontecimiento tremendamente improbable, un
acontecimiento con una probabilidad tan pequeña como para tener
esencialmente el valor de cero.
Aquí
este evolucionista da por supuesto que la vida surgió por
procesos naturales fisicoquímicos sin intervención
sobrenatural, y que se debe hallar una explicación a ello sobre
esta base. Thomas Huxley, el famoso evolucionista y defensor de Darwin,
afirmó en su día:
Queda claro que la doctrina de la Evolución es
directamente antagónica a la de la Creación. La
Evolución, si es aceptada coherentemente, hace imposible creer
la Biblia.
SOLAMENTE UNA
PEQUEÑA PARTE DEL PROBLEMA
Se
debe insistir que en estos casos se suponen condiciones enormemente
favorables para el origen de la vida al azar: (1) Se da por supuesto
que sólo están presentes los 20 alfa-aminoácidos;
(2) que de estos 20 alfa-aminoácidos 19 son todos de la variedad
L- (levógira) (uno de ellos, la glicina, tiene una
estructura tal que su imagen en el espejo es ella misma); (3) se
permiten concentraciones elevadas de reactivos absolutamente
impensables desde un punto de vista termodinámico, pues no
habrían podido acumularse jamás en las concentraciones
empleadas en los cálculos; (4) se emplean unas enormes
velocidades de reacción; (5) se dan 10.000 millones de
años para este proceso cuando, desde un punto de vista
evolucionista se habría dispuesto como máximo de
alrededor de 1.000 millones.
Por
otra parte, en los cálculos no entran cuestiones como (1) el
problema del sol débil. Se trata del problema de que
desde una perspectiva cosmogónica evolucionista, que afirma el
origen del universo por evolución estelar, la luminosidad del
sol habría sido alrededor del 60 por ciento de la actual hace 4
mil millones de años, con la consecuencia de que la tierra
habría sido un planeta helado [Philip Handler, ed., Biology
and the Future of Man (New York: Oxford University Press, 1970);
pág. 174]: en base de modelos climáticos, con un 1 por
ciento menos de radiación solar se padecería una edad
glacial no demasiado severa; pero si el sol tuviera una luminosidad del
75 al 80 por ciento de la actual, la tierra debería quedar
cubierta de hielo [John A. Eddy, en «It's About Time», Geotimes,
Sept. 1978, pág. 18]. (2) La presencia de especies
químicas hostiles a la concatenación de
aminoácidos en cadenas peptídicas (la presencia de
azúcares o aldehidos, por ejemplo, debido a su gran afinidad con
los aminoácidos, llevaría a una reacción irreversible
(reacción de Maillard) que provocaría la precipitación
de unos compuestos estables macromoleculares insolubles. De esta
manera, con azúcares o aldehidos en exceso,
desaparecerían virtualmente todos los aminoácidos
del medio reactivo; a la inversa, con los aminoácidos en exceso,
desaparecerían virtualmente todos los azúcares,
compuestos esenciales de las cadenas del ADN y ARN. (3) También
desde la perspectiva de la formación del ADN y ARN, y de otros
compuestos biológicos de suma importancia, como el ADP y ATP, la
presencia de cationes como el Ca[2+] y el Mg[2+] en el «mar
primordial» provocaría asimismo la precipitación
del fósforo (P) presente, impidiendo su inserción en el
proceso, y por ello imposibilitando la formación de los
compuestos ya mencionados, de los que el fósforo es un vital
elemento. (4) No se ha tocado la importantísima cuestión
de la necesaria presencia, en un proceso natural, de
isómeros ópticos indistintamente levógiros
y dextrógiros, factor éste que eleva de una
manera enorme el grado de imposibilidad de una concatenación de
cadenas peptídicas biológicamente viables por medios
aleatorios.
En
resumidas cuentas, a la hora de emplear el cálculo de
probabilidades del proceso, se han admitido una serie de condiciones
favorables que permiten la marcha del proceso que se calcula. En
realidad, hay una gran cantidad de consideraciones, de cariz
cosmológico, físico y químico, que se han
mencionado de pasada, y que impedirían el mismo
arranque del proceso. No sólo nos encontramos con un
billete de lotería con una imposibilidad supercósmica de
salir premiado, sino que, aún peor, no nos es posible
comprarlo.
Se
debe remarcar, pues, por las razones expuestas y muchas más en
las que no es posible entrar aquí ahora, que los cálculos
empleados han sido extremadamente generosos para la postura
naturalista, y que en realidad los números que expresan la
imposibilidad son de grado supercósmico por un gran margen.
HAGA JUEGO A SU PROPIO
RIESGO
Ningún
jugador arriesgaría toda su fortuna si la probabilidad fuera de
10 a 1 contra él. Ningún hombre de negocios
asumiría un riesgo si, una vez que fueran considerados todos los
factores, la posibilidad de éxito fuera sólo de 1 a 10.
La posibilidad de que la vida se originara a través de un
proceso puramente materialista es realmente nula, y ello por un margen
que, a todos los efectos, demuestra su total imposibilidad. Por lo
tanto, la conclusión es que la vida tuvo que ser originada por
medio de una acción inteligente. Cualquier otra
conclusión se enfrenta contra un número abrumador que
expresa imposibilidad.
LA VERDAD Y LAS
CONSECUENCIAS
Se
debe concluir que, sin excepción alguna, la vida se ha
transmitido de padres a hijos en una cadena que arranca de la
creación de cada tipo viviente. Y que la vida fue originada por
el meticuloso designio de un Creador infinitamente inteligente.
BIBLIOGRAFÍA
Berhnard,
Sydney, Estructura y Función de los Enzimas, H. Blume
Ed. (Madrid 1977).
Borel, Émile, Las Probabilidades y la vida Ed. Orbis
(Barcelona, 1986).
Gish, Duane T., Especulaciones y experimentos relacionados con
teorías sobre el origen de la vida: Crítica, CLIE
(Terrassa, 1981).
Salet, George, Azar y Certeza, Ed. Alhambra, Col. Exedra n(a)
95 (Madrid, 1975).
FUENTE: Basado en el
artículo «The Origin of the First Living System»,
del doctor William E. James, en el Simposio A Challenge to Biology. ©
Copyright 1977 por Bible-Science Ass., Inc. Revisado y ampliado por
Santiago Escuain.
LOS AUTORES
El doctor William E. James es investigador
médico, y ha colaborado en diversos proyectos relacionados con
los programas Titán y Apolo en la empresa Martin Marietta Corp. Vuelve al encabezamiento
Santiago Escuain es químico, habiendo cursado
los estudios de Ingeniería Técnica en esta rama, y con
muchos años de experiencia en química aplicada. Vuelve al encabezamiento
 Arriba un nivel Arriba un nivel
De vuelta al índice general
De vuelta a la página principal
©
Por la adaptación: Santiago Escuain 1984
© Copyright SEDIN 1997 para el formato electrónico -
www.sedin.org. Este texto se puede reproducir
libremente para fines no comerciales y citando la procedencia y
dirección de SEDIN, así como esta nota en su integridad.
-
SEDIN
Servicio Evangélico - Documentación - Información
Apartat 2002
08200 SABADELL
(Barcelona) ESPAÑA
|
Lecturas complementarias
David Coppedge
El
naufragio en las investigaciones sobre el origen de la vida (2007)
Dean H. Kenyon, Ph.D.
Más allá de la mentalidad
naturalista en las investigaciones sobre el origen de la vida (1989)
Nancy Pearcey
El
primer paso
La Evolución Química -
Entrevista con el doctor Charles B. Thaxton - Coautor de The Mystery of
Life’s Origin
(1985)
Duane
T. Gish, Ph.D.
Especulaciones y
experimentos sobre el Origen de la vida: Crítica (1972)
Visite también:
El Museo de Máquinas Moleculares
Índice de
boletines
Índice de línea
sobre línea
Página principal
Índice general
castellano
Libros recomendados
orígenes
vida cristiana
bibliografía general
Coordinadora Creacionista
Museo de Máquinas
Moleculares
Temas de actualidad
Documentos en PDF
(clasificados por temas)




|