Dean H.
Kenyon
Más
allá de la mentalidad naturalista en las investigaciones
sobre el origen de la vida
Introducción
Si se acepta la concepción evolucionista general
de la historia de la vida sobre la tierra, no es difícil creer
que la vida debe haber surgido de manera natural1 por
medio de evolución química. Si la vida se ramifico de
modo natural a partir de organismos de una sola célula hasta la
maravillosa complejidad de las formas de vida que tenemos en la
actualidad, ¿por qué no pudieron los primeros paquetes
mínimos de información genética (precisos para
sustentar la vida) haber surgido de manera espontánea como
resultado de las propiedades de la materia carbonacea? Y desde luego,
si uno tiene esta mentalidad, es muy difícil poder ver
cómo el origen de la vida puede haber sido por otra vía
que por la de evolución química.
Desde
un punto de vista naturalista, uno considera su investigación
como un esfuerzo para encontrar caminos químicos posibles par el
surgimiento de la vida. El objetivo es limitar el conjunto de caminos
posibles y producir al menos un bosquejo de la transición de la
no-vida a la vida. Es de suponer que muchos procesos químicos
que tuvieron lugar en la superficie del globo primigenio serán
parcialmente duplicados en las simulaciones de laboratorio.
Si,
sin embargo, uno abriga dudas acerca de la perspectiva evolutiva
general del origen de la vida y mantiene abierta la posibilidad de que alguna
otra explicación pudiera ser correcta, queda en libertad
para enfocar el problema del origen de la vida desde un ángulo
distinto. Una manera de enunciar las presuposiciones de esta pesona
sería ésta: La vida puede que haya surgido
espontáneamente (de manera natural) en este planeta, o puede que
no. Por causa de la necesidad metodológica (Kenyon y
Steinman, 1969, pág. 31; Kenyon 1984a) uno podría suponer
inicialmente que la vida surgió de manera natural. Podría
entonces tratar de simular acontecimientos químicos y
físicos que pudieran haber estado envueltos en un proceso
así en la Tierra primitiva. Si aconteciera que algunos de los
resultados de nuestros experimentos no son congruentes con la
presuposición de un origen natural, entonces evaluaría de
manera cuidadosa estos resultados a la luz de la posibilidad de que
dieran apoyo a otra perspectiva general. Además, al interpretar
los resultados de cualquier experimento de simulación en
particular, uno debería tener cuidadosamente en mente los
resultados de otros experimentos de este tipo, así como
cualesquiera hechos relevantes químicos y físicos —
incluyendo cualquier evidencia geoquímica que tuviera que ver
con las condiciones de la simulación. En otras palabras, uno
entretendría de manera seria la posibilidad de que los
resultados experimentales refutaran la misma suposición de
evolución química. Desde luego, hay lugar para esta
apertura: no podemos tener la misma certidumbre acerca de nuestras
explicaciones para el origen de la vida que la que pudiéramos
esperar en «ciencia operativa», como han argumentado
persuasivamente Thaxton et al. (1984, págs. 202-206).
Los experimentos de Miller
Algunos
aspectos de la obra de Stanley Miller acerca del origen de la vida nos
serán de ayuda para comparar las dos perspectivas generales
anteriormente bosquejadas. Stanley Miller ha sido uno de los pioneros
en estudios experimentales del origen de la vida. Él
llevó los primeros experimentos sistemáticos de
simulación hace más de 35 años (Miller, 1953).
Desde entonces ha producido un cuerpo de investigación adicional
(Miller, 1955, 1957; Ring et al.,
1972; Ring y Miller, 1984) a lo largo de las líneas de su obra
original: simulaciones de atmósfera primitiva empleando descarga
eléctrica como fuente de energía. También ha
escrito síntesis de la situación del campo de la
«evolución química prebiótica» (Miller
y Orgel, 1974; Miller, 1984). La obra de Miller ciertamente satisface
el fructífero criterio de la buena investigación
científica. Además, su método de «simular la
totalidad» es quizá el enfoque más razonable para
hacer estudios «de simulación».
Si uno desea saber si pudo haber tenido lugar un
origen evolutivo químico de la vida, no puede simplemente dar
por supuesto que sí tuvo lugar, y luego argumentar que las
condiciones sobre la Tierra prebiótica tuvieron que ser tales
que permitieran que esto tuviera lugar.
En
sus artículos, Miller es precavido, y tiende a apegarse a los
verdaderos datos de laboratorio cuando evalúa los resultados de
los experimentos de evolución química. No es persona dada
a hacer pretensiones excesivas acerca de progreso en el logro de
posibles caminos prebióticos, y yo aprecio en gran manera su
actitud realista. Contrasta de una manera refrescante con las
extravagantes conclusiones de otros experimentadores. Aunque Miller
cree que se ha hecho un progreso razonable acerca de posibles caminos
de síntesis de biomonómeros prebióticos, y que se
ha logrado un modesto progreso en al menos los inicios de
síntesis de biopolímeros, reconoce que es
poquísimo lo que se «conoce» más allá
de esto.
Hasta aquí todo está muy bien, pero hay
más. Miller, como los otros evolucionistas químicos,
comienza dando por supuesto que algún proceso naturalista dio
origen al primer ser vivo. Luego, arguyendo en base de las propiedades
de compuestos orgánicos y de los resultados de experimentos de
simulación, hace varias inferencias acerca de las condiciones en
la superficie de la tierra primitiva. Por ejemplo, los compuestos
orgánicos se descomponen en presencia de O2;
por ello, el O2
tiene que haber estado prácticamente ausente de la
atmósfera primitiva (Miller, 1984). Se mencionan otras razones
para creer que la primitiva atmósfera de la tierra
contenía muy poco O2, pero ninguna
de ellas parece tan convincente como la acabada de citar.2
Si
se acepta la posibilidad de que la vida se originara por medios no
naturalistas, es apropiado razonar de manera diferente acerca de la
cuestión del O2 en la
atmósfera primitiva: la atmósfera primitiva puede haber o
no haber contenido cantidades sustanciales de O2.
Pero esta cuestión debe decidirse en base de evidencia
independiente, no en base de los resultados de experimentos de
simulación ni de las necesidades de la hipótesis de
Haldane/Oparín (Kenyon, 1984b). Si uno desea saber si pudo haber
tenido lugar un origen evolutivo químico de la vida, no puede
simplemente dar por supuesto que sí tuvo lugar, y luego
argumentar que las condiciones sobre la Tierra prebiótica
tuvieron que ser tales que permitieran que esto tuviera lugar.
Al
tratar de la temperatura de la Tierra primitiva, Miller (1984) dice lo
siguiente:
La temperatura del océano primitivo no se conoce,
pero se puede decir que la inestabilidad de varios compuestos y
polímeros orgánicos constituye un argumento convincente
de que la vida no pudo haber surgido en los océanos a no ser que
la temperatura estuviera por debajo de los 25°C.
Una
vez más se emplean aquí las necesidades de un origen
evolutivo químico de la vida para limitar las condiciones
primitivas de la superficie de la tierra. Pero, ¿no sería
más razonable intentar sustentar un argumento acerca de la
temperatura de los océanos primitivos con independencia de estas
necesidades? Si no tenemos a mano la información necesaria para
dar respuesta a esta pregunta, deberíamos suspender nuestro
juicio.
Miller
se muestra precavido acerca del tema de la composición de la
atmósfera primitiva, pero se hace evidente la misma clase de
razonamiento:
Alguna de la química orgánica [esto es, de los
experimentos de simulación] hace predicciones explícitas
acerca de los constituyentes de la atmósfera. Tales
consideraciones no pueden demostrar que la tierra tuvo una determinada
atmósfera primitiva, pero los condicionantes de la
síntesis prebiótica debería ser una
consideración principal. (Miller, 1984, énfasis
añadido).
Aquí
Miller se refiere al hecho experimental de que cuanto más
reductora sea la atmósfera simulada (p.e., CH4,
NH3, H2O
frente a CO2, N2,
H2O) tanto mayor la producción de
compuestos orgánicos como los aminoácidos, los
constituyentes básicos de las proteínas. Se considera que
los resultados de los experimentos de simulación aportan un
poderoso apoyo a la postura de que la atmósfera primitiva de la
Tierra era reductora. Pero, una vez más, tal razonamiento es
persuasivo sólo si uno da por supuesto que tiene que haberse
dado una evolución química.
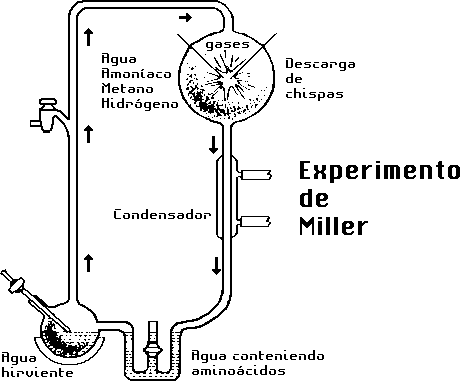
Aparato empleado en los experimentos de Miller de descargas
eléctricas para conseguir aminoácidos.
(Según R. Jastrow, 1967, Red Giants and White Dwarfs.
Nueva York: Harper and Row, pág. 134.)
Abundan ejemplos de este tipo de razonamiento, pero será
suficiente con dos más (Miller, 1984). Por cuanto los estudios
de laboratorio indican que es necesario un cierto nivel de NH4+
(alrededor de 0,01 M, que se corresponde con una presión parcial
atmosférica de NH3 de alrededor de
4 x 10E6 atm a 25° C) para una síntesis eficaz de
aminoácidos, se supone que al menos esta cantidad de NH4+
tuvo que haber estado presente en los océanos primitivos. En
otros experimentos se mostró que los ribósidos de
pirimidinas (componentes del ARN) no se forman en mezclas calentadas de
ribosa y pirimidinas (mientras que las mezclas calentadas de ribosa y
de purinas sí que dan ribósidos de purinas). Miller ve
dos optativas aquí: 1) Quizá no se probaron las
condiciones adecuadas, ó, 2) no hubo ribósidos de
pirimidinas en los primeros organismos. La optativa 1 es posible,
naturalmente, pero la optativa 2 exhibe la misma falacia argumental que
nuestros primeros ejemplos. ¿Cómo pueden unos resultados
negativos sugerir que un compuesto determinado no fue necesario para el
origen de la vida? Una conclusión así sólo tiene
sentido en base de la previa suposición de que la
evolución química ha tenido que ocurrir, y que la
«simulación» en cuestión duplica de una
manera razonable los acontecimientos prebióticos en la Tierra
primitiva.
Vale
la pena enfatizar que la mayoría de los que están
acutalmente llevando a cabo experimentos acerca del origen de la vida,
si no todos, no sólo dan por supuesto que tuvo lugar un proceso
evolutivo, sino que además consideran que están indagando
los detalles de este pretendido proceso en lugar de estar generando un
cuerpo de datos de laboratorio que, cuando sea añadido al
conocimiento químico y físico ya existente, podría
ayudar a decidir si la evolución química tuvo lugar o
no tuvo lugar.
Los experimentos de Fox
Sidney
Fox es otro experimentador que ha generado un cuerpo impresionante de
investigación experimental sobre el origen de la vida (Fox,
1956, 1964, 1974, 1978, 1981; Fox y Dose, 1977; Fox y Harada, 1960; Fox
y Nakashima, 1980; Fox y Yuyama, 1964). Su perspectiva básica
parece ser algo como lo que sigue: El origen de la vida fue
indudablemente por medios naturalistas. Los caminos más
probables son los que se presentan en la teoría de origen por
proteinoides, y hay un extenso sustento experimental para este punto de
vista. La teoría abarca todo la gama de los eventos
prebióticos, desde los gases primitivos hasta microsistemas
orgánicos. El enfoque de Fox ha sido enormemente
fructífero en la generación de experimentos, y prosigue
la investigación a lo largo de estas líneas en muchos
laboratorios por todo el mundo (Matsuno et al., eds., 1984).
El
enfoque de Fox es muy diferente al de Miller. Miller acentúa la
«simulación global». Esto es, él recrea lo
que se supone fue la atmósfera primitiva, y la provee con
posibles fuentes prebióticas de energía. No hay
interferencia del investigador hasta que se analizan los productos del
experimento. La marcha química de estos experimentos es bastante
compleja, y aunque se producen muchos compuestos biológicos y no
biológicos, de bajo peso molecular, la tendencia dominante de
los procesos químicos es hacia la producción de material
macromolecular no biológico (Folsome, 1979; Folsome et at.,
1975; Kenyon, 1984b). Además, no se encuentran en estos
experimentos ni proteínas ni ácidos nucleicos, y los
aminoácidos son todos racémicos de una manera precisa,
condición ésta que difiere marcadamente de la presencia
exclusiva de L-aminoácidos en las proteínas de los
organismos vivientes.
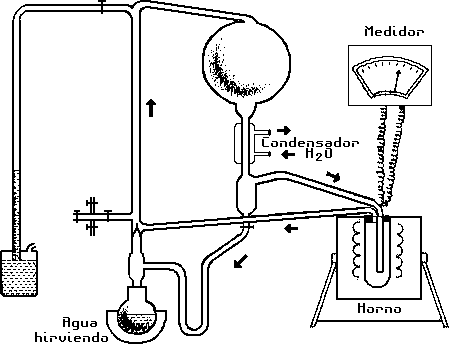
Aparato empleado para la síntesis termal de
aminoácidos a partir de gases simples.
(Según Harada y Fox, 1965, en Origins of Prebiotic Systems.
Nueva York: Academic Press, pág. 187.)
Fox,
en cambio, destaca el papel prebiótico de una clase de
biomonómero, esto es, de los aminoácidos. En los
experimentos de formación de proteinoides emplea él
sólo aminoácidos (por lo general la forma L) como
reactivos iniciales. Una simplificación tan artificial del
sistema reactivo cae de inmediato bajo sospechas. ¿Por
qué no habría habido presentes también muchos
otros tipos de compuestos junto con los aminoácidos? Algunos de
estos, especialmente aldehidos y azúcares, habrían desde
luego reaccionado en reacción cruzada con aminoácidos
(vía la reacción de Maillard, Ellis, 1959) para formar
material macromolecular insoluble no biológico en lugar de
proteinoides (Kenyon, 1984a). ¿Por qué, entonces, no se
hacen experimentos en los que se añadan aldehidos y/o
azúcares a los aminoácidos? ¿Quizá porque
con toda certeza se generaría material no biológico
intratable, y se desalentaría la futura experimentación?
Evidentemente, Fox da por supuesto que las intensas reacciones cruzadas
de interferencia, como las que aparecen en los experimentos de Miller,
no fueron importantes en la evolución química
prebiótica, a pesar de que se sabe que la reacción de
Maillard tiene lugar en un margen muy amplio de condiciones de
reacción (Ellis, 1959; Nissenbaum et al., 1975).
La
anterior discusión de los problemas en enfoques empíricos
actuales acerca del origen de la vida, naturalmente, deja de ser
completa. Para análisis más extensos de fallos en la
investigación sobre evolución química, se remite
al lector a Kenyon (1984a, 1984b) y a Thaxton et al. (1984).
La atmósfera primitiva puede haber o no haber
contenido cantidades sustanciales de O2. Pero esta cuestión
debe decidirse en base de evidencia independiente, no en base de los
resultados de experimentos de simulación ni de las necesidades
de la hipótesis de Haldane/Oparín.
El origen de la información genética y del
sistema de síntesis de proteínas
Desde
mi punto de vista, el mayor reto a cualquier teoría naturalista
de biogénesis es el problema del origen de la información
genética. Este problema, enunciado con brevedad, es como sigue:
¿Cómo unas secuencias específicas de bases
conteniendo una información biológicamente relevante
se acumularon de manera espontánea (naturalísticamente)
en polinucleótidos sobre la Tierra prebiótica? Estas
secuencias tienen que haber llegado a abarcar todas las necesidades de
la célula mínima, y, por tanto, todas tienen que haberse
juntado dentro del mismo pequeñísimo volumen.
El
problema conceptual es abrumador. Para que la información
biológica funcione en el estado de vida, tiene que ser traducida
a las secuencias de aminoácidos de al menos varios cientos y
posiblemente varios miles de diferentes enzimas y otras
proteínas. La maquinaria molecular para la duplicación,
transcripción y traducción de la información
genética (y al menos se precisa de 70 proteínas
específicas sólo para estas funciones) tiene que estar ya
toda en funcionamiento para que el sistema efectúe su ciclo
completo de funciones la primera vez. Considerando la cantidad total de
secuencias posibles de las bases en un polidesoxiribonucleótido
lo suficientemente largo para codificar todos los enzimas y otras
proteínas necesarias para el estado de vida (Ambrose, 1982,
pág. 135), la probabilidad de formar incluso una pequeña
fracción de la información necesaria por procesos
naturalistas —incluso a lo largo de períodos miles de veces
más largos que toda la duración que se supone al cosmos—
es virtualmente nula (Eden, 1967; Hoyle y Wickramasinghe, 1981,
págs. 23-33). Las minúsculas indicaciones experimentales
de ordenación en secuencias no aleatorias ya sea en
polinucleótidos, ya en polipéptidos (Fox, 1978) en
estudios de «simulación» son totalmente inadecuados
para afrontar este problema (cf. Yockey, 1977, 1981). El problema no es
sencillamente la demostración de alguna ordenación no
aleatoria, sino la de demostrar una ordenación biológicamente
relevante.
Luego,
naturalmente, existen los problemas previos de la acumulación
espontánea de elementos constitutivos de los ácidos
nucleicos (nucleótidos), y la incorporación de
sólo los nucleótidos naturales en moléculas
primitivas de ácidos nucleicos. Shapiro (1984) ha presentado un
convincente argumento de que tales acontecimientos habrían sido
enormemente improbables en la Tierra prebiótica.
Algunos
científicos mantienen que ya se ha logrado un significativo
progreso experimental acerca del problema del origen de la
información genética. Yo clasificaría a Fox entre
los que pertenecen a este grupo. Su creencia es que una mayor
investigación acerca del modelo del proteinoide dará
finalmente la respuesta: Puede que se precise de tiempo, pero ya
tenemos claves experimentales indicando el camino (Fox, 1981). Los
proteinoides mismos fueron los primeros portadores significativos de
información biológica.
Otros
investigadores han destacado la enorme dificultad del problema,
respondiendo con una variedad de propuestas y posiciones diferentes.
Kuppers
(1983, pág. 279) declara que «La probabilidad de que un
sistema genéticamente integrado se monte espontáneamente
por pura casualidad es insignificantemente pequeña. Esto es
cierto incluso de la síntesis al azar de una pequeña
molécula de enzima optimizada para alguna función
biológica». Tanto Kuppers (1983) como Eigen (1971; Eigen et
al.,
1981) han respondido a este reto desarrollando unas (similares)
complicadas teorías acerca del origen naturalista de la
información genética basadas en el concepto del
«hiperciclo». Se conciben los primitivos hiperciclos como
agrupaciones acopladas de moléculas de ARN auto-duplicantes y
polipéptidos catalizadores asociados. Se afirma que estos
sistemas químicos, al principio relativamente sencillos, tienen
la propiedad de evolucionar espontáneamente hacia sistemas
moleculares genéticos integrados, esto es, las primeras
células vivas.
Por
atractivas que puedan ser estas ideas de Eigen y Kuppers, el sustento
experimental de las mismas es desde luego bien parco. Además,
esta línea de pensamiento parece adolecer de un fallo fatal en
cuanto a que no puede explicar el origen del ARN y de las
proteínas catalíticas que necesita para el surgimiento de
los hiperciclos. Eigen y Kuppers dan por supuesto que los experimentos
de evolución química han mostrado cómo tales
moléculas pudieron haber surgido en la Tierra primitiva, pero la
verdad es que no existe ninguna evidencia que sustente tal tesis. Toda
la información química de que disponemos arguye
intensamente en contra de una aparición prebiótica
espontánea de ácidos nucleicos (Shapiro, 1984). Admiro
los esfuerzos que estos experimentadores han hecho y que creen que
deben continuar, pero dudo de que sus presentes perspectivas tengan
mucho que ver con el verdadero origen de la vida.
Cairns-Smith
(1982) también reconoce claramente la seriedad del problema. El
actual sistema genético molecular no se pudo desarrollar
espontáneamente en la Tierra primitiva. ¿Brotó
entonces la primera vida de manera naturalista? Sí, pero los
primeros organismos eran sistemas inorgánicos basados en
minerales de arcilla. Posteriormente, el sistema carbonáceo
ocupó el campo. Ésta es una idea novedosa, y merece
más estudio por parte tanto de creacionistas como de
evolucionistas químicos. Hay posibilidades aquí para
experimentos significativos.
Monod
(1972) argumentaba a lo largo de las siguientes líneas
básicas. El origen naturalista del primer sistema
genético era sumamente improbable. Pero estamos aquí y el
origen de la vida tiene que haber sido un acontecimiento o proceso
naturalista. ¿Qué otra cosa pudo ser? Por ello, el origen
de la vida fue de hecho un acontecimiento (o conjunto de
acontecimientos) muy raro, de baja probabilidad. Monod pudiera estar en
lo cierto, pero no tenemos la posibilidad de demostrar la
corrección de su postura, porque estamos tratando aquí
con lo que viene a ser un «milagro naturalista».
Naturalmente, la conclusión del creacionista de que la primera
información genética provino de un origen sobrenatural
tampoco está sujeta a una verificación experimental
directa.
Moviéndonos
a lo largo de nuestra gama de posturas acerca del problema del origen
de la información genética llegamos a Hubert Yockey,
aunque podría ser situado algo más adelante. Yockey
parece considerar totalmente inadecuados todos los escenarios
naturalistas propuestos hasta ahora para poder dar cuenta de ninguna
cantidad significativa de información genética o de
información biológica almacenada en polipéptidos
(Yockey, 1981). Como mucho, ve un cierto potencial para producir una
pequeña proteína aislada. Sus cálculos
probabilísticos tienen en cuenta el hecho de que hay 39
isómeros ópticos de los 20 aminoácidos
constituyentes de las proteínas, punto éste no destacado
por otros autores (Yockey, 1977).
Crick
reconoce el problema de un origen naturalista de la vida en la Tierra
primitiva, pero no lo descarta. Sugiere él la posibilidad
alternativa de que la primera vida (microbiana) en la Tierra puede
haber llegado de otras partes en el cosmos, quizá transportada
en una nave espacial no tripulada (Crick, 1981, págs. 141-153).
Esta solución, llamada «panspermia dirigida», nos
deja con el enojoso problema de cómo surgió la vida en el
planeta desde el que fueron enviados los microbios. Quizá fue
por panspermia dirigida, y así ad infinitum.
Hoyle
y Wickramasinghe, en una notable serie de libros (p.e., Hoyle, 1983;
Hoyle y Wickramasinghe, 1981), han desarollado la postura de que la
vida no pudo surgir espontáneamente (esto es, al azar) en
ningún lugar del cosmos. Alguna clase de inteligencia tiene que
haber conducido el proceso. Pero no queda claro del todo cómo
debemos concebir esta inteligencia. En el libro El Universo
Inteligente (1983, pág. 136) Hoyle dice: «La
inteligencia responsable de la creación de la vida basada en el
carbón en la teoría cósmica queda firmemente dentro
del Universo y es subordinada a él». Él distingue
de manera clara esta inteligencia inmanente del concepto de un Dios
sobrenatural creador fuera del espacio y del tiempo.
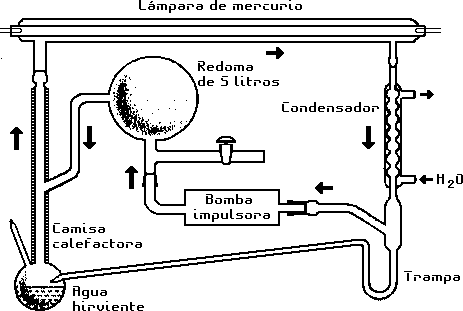
Aparato empleado para la síntesis de aminoácidos mediante
rayos ultravioleta de lámpara de mercurio.
(Según Kenyon y Steinman, 1969. Biochemical Predestination,
Nueva York: McGraw-Hill, pág. 135.)
Mi propia postura acerca de este tema se puede bosquejar de la
siguiente manera: No creo que el sistema de codificación
genética y de síntesis de proteínas pudo haberse
desarrollado de manera naturalista a partir de configuraciones
materiales más simples. Cuanto más aprendemos acerca de
los detalles moleculares del sistema, tanto más fuerte se hace
esta posición. Cuando se toman en cuenta todas las líneas
pertinentes de evidencia y se afrontan de manera franca todos los
problemas, creo que debemos llegar a la conclusión de que la
vida debe su origen a una fuente externa a la naturaleza (Kenyon,
1984b). Algunas de las principales líneas de evidencia y de
razonamiento que sustentan esta postura son como siguen:
(1) La virtual imposibilidad de un origen espontáneo
de la información genética, incluso en 10 o 20 mil
millones de años.
(2) El hecho de que la tendencia dominante en las
«simulaciones del total» es hacia la producción de
materiales macromoleculares no biológicos intratables (Folsome,
1979; Folsome et al., 1975; Kenyon, 1984b). En estos
experimentos no se encuentran biopolímeros. Predominan las
reacciones cruzadas interferidoras en las simulaciones más
«realistas».
(3) Los experimentos de simulación producen
rutinariamente mezclas racémicas de compuestos orgánicos
(Dickerson, 1978; Folsome, 1979). Todos los intentos experimentales de
encontrar una base naturalista plausible par la acumulación
preferencial primigenia de L-aminoácidos han fracasado (Bonner,
1972; Bonner et al., 1980).
(4) La evidencia geoquímica es congruente con la
conclusión de que hubo cantidades significativas de O2
en la primitiva atmósfera de la tierra (Clemmey y Badham, 1982;
Dimroth y Kimberley, 1976). Este O2
habría eliminado cualquier pretendida evolución
química en sus más primitivos estadios.
Finalmente,
llegamos a A.E. Wilder-Smith, que desde luego reconoce el problema del
origen de la información genética (Wilder-Smith, 1970).
Él observa que los experimentos de
«simulación» ideados para hallar posibles caminos
prebióticos de síntesis de componentes del sistema
genético molecular son a menudo sumamente complejos y que se
apartan significativamente de las condiciones prebióticas
«razonables». Por ejemplo, las mezclas reactivas iniciales
son artificialmente simplificadas para asegurar la formación de
productos de interés. No estamos viendo en ello lo que pueda
hacer una materia carbonácea relativamente carente de ayudas con
un aporte de energía. Más bien estamos manipulando de
manera deliberada las condiciones experimentales para poder tener un
control significativo sobre el resultado (cf. Thaxton et al.,
1984, págs. 104-110).
También es evidente un control inapropiado en los
experimentos de proteinoides de Fox, que comienzan con mezclas
reactivas simplificadas artificialmente: esto es: sólo con
aminoácidos. Se podría decir que cuanto más
interferimos de esta forma aplicando «tecnología»
química, tanto más nuestras actividades (aunque muy
remotamente) se asemejan a las de alguna agencia sobrenatural que trajo
a la existencia la primera vida. Los reactivos químicos dejados
a sí mismos (esto es, en ausencia de una información
genética preexistente) no llegan a la vida. Se precisa de una
manipulación externa conductora.3 Es a esto a
lo que señala toda nuestra evidencia experimental, y es desde
luego la conclusión del mejor razonamiento teórico.
Mantener la creencia en una biogénesis naturalista, sabiendo lo
que sabemos acerca de las propiedades de la materia y de la
energía, equivale « ... simplemente a arrojar la toalla
científica» (Wilder-Smith, 1981, pág. 146).
Conclusiones
Es
difícil comprender por qué unos científicos
están dispuestos a admitir el pensamiento de que una
inteligencia de alguna clase (dentro o fuera del universo) estuvo
involucrada en la aparición de la primera vida, mientras que
otros aparentemente no lo están, y aquí no voy a proponer
una explicación de ello. Pero lo que queda ilustrado con la gama
de puntos de vista descritos en la anterior sección es que hay
un margen de posiciones intermedias que cubren la distancia intelectual
entre investigadores como Fox o Miller, por una parte, y otros como
Hoyle, Kenyon o Wilder-Smith por la otra. Por ello, no es necesario
salir del mundo racional para dirigirse en dirección de una
comprensión científica francamente
«teísta» del origen de la vida, a no ser,
naturalmente, que uno considere cualquier desviación del
«optimismo naturalista» como irracional y anti-intelectual.
La
conclusión de que la vida demanda una creación
sobrenatural (o al menos una inteligencia creadora dentro del universo)
les parecerá prematura a muchos, incluso si se conceden algunos
méritos a las principales críticas y argumentaciones
positivas creacionistas. Y seguirá habiendo los que
querrán proseguir con sus actuales enfoques, sin importar lo que
muestren los resultados. Siempre podrán decir:
«Todavía no hemos encontrado las condiciones
experimentales correctas». O, «Sólo hemos estado
haciendo experimentos específicamente relacionados con el
problema del origen de la vida durante 30 años. Dadnos tiempo. A
fin de cuentas, se trata de un problema muy difícil. Al final
encontraremos una descripción evolucionista química
plausible». O, «Es muy poco lo que se conoce actualmente
del sistema genético molecular».
Consideremos
lo que podría suceder a las tendencias en la
investigación del origen de la vida si de repente todos los
miembros del ISSOL (International Society for Study of the Origin of
Life - Sociedad Internacional para el Estudio del Origen de la Vida)
decidieran considerar seriamentee la posibilidad de que la vida hubiera
sido creada por un agente inteligente. Supongamos también (para
combinar lo ridículo con lo absurdo) que decidieran probar y
asumir una actitud mental en la que verdaderamente no estuvieran
entregados ni a la postura evolucionista ni a la creacionista. Por
extraña que pueda parecer esta sugerencia, me parece que
constituye exactamente el tipo de nuevo principio que necesitamos para
el problema del origen de la vida en la actualidad.
Como
mínimo, una reorientación de pensamiento de este tipo
generaría probablemente unas nuevas discusiones de
carácter notable. Por ejemplo, podrían aparecer listas
como ésta en las pizarras de los laboratorios:
(1) No hay demostración de transición de lo
inanimado a materia viviente en ausencia de sustancias preexistentes
portadoras de información genética.
(2) No hay demostración en laboratorio de estados
precursores realistas.
(3) No hay ninguna descripción teórica
convincente de posibles estados precursores.
(4) No hay registro fósil de estados precursores,
incluyendo la inexistencia de evidencia geoquímica de un
«caldo orgánico prebiótico» (Corliss et al.,
1981; Nissenbaum et al., 1975; Thaxton et al., 1984,
pág. 57).
(5) No hay neobiogénesis (origen de la vida hoy
día).
(6) No hay evidencia de vida extraterrestre ni de estados
materiales precursores encontrados en la exploración espacial
hasta el día de hoy.
En
base de una lista así, se podría pasar a considerar
cuestiones como las siguientes: ¿Es posible mostrar
experimentalmente que la materia y la energía no pueden
organizarse por sí mismas en un sistema vivo, si no hay materia
viva ya presente? El fracaso en demostrar la transición no
constituye prueba suficiente, desde luego, para excluir la posibilidad.
¿Existen criterios formales útiles que nos puedan servir
de ayuda para determinar si algún sistema natural recibió
su diseño impuesto desde fuera, o si el orden se generó
de manera natural y espontánea? ¿Es el sistema
genético molecular en realidad a este respecto como un sistema
complejo manufacturado, como afirman los creacionistas? (para un
tratamiento preliminar de esta cuestión, véase Parker,
1980, págs. 2-15). Si es así, ¿cuáles son
las implicaciones de tal conclusión para la planificación
de experimentos acerca del origen de los sistemas de
codificación genética y de síntesis de
proteínas?
La
discusión a lo largo de estas líneas llevaría
naturalmente a las siguientes cuestiones de mayor entidad:
¿Qué sucedería a la investigación sobre el
origen de la vida en el laboratorio en que uno trabaja, si se llega a
aceptar la tesis creacionista de los orígenes biológicos
(Kenyon 1984b)? ¿Cesaría del todo? ¿No
quedaría uno entonces limitado, por lo que respecta a las tareas
experimentales, a problemas de ciencia operativa (Thaxton et al.,
1984, págs. 8, 204)? Si todos aceptáramos este punto de
vista, ¿qué sucedería con nuestras publicaciones y
nuestras carreras?
Quizá
sería más prudente comenzar con una reorientación
más limitada del pensamiento. Quizá algunos
equipos de investigación podrían tener a un
científico creacionista, o al menos a alguien abierto a la
posibilidad de una explicación no naturalista de los
orígenes, que tuviera parte en la planificación de los
experimentos. Se podría dar la sugerencia de modificar algunos
de los experimentos de Fox añadiendo, por ejemplo,
aldehídos y azúcares. O podrían repetirse los
experimentos de Miller con cantidades significativas de O2
presentes en el aparato, con la precaución de que no haya H2 en
la mezcla inicial de gases, para impedir una explosión.4 De esta manera, el trabajo experimental podría
continuar muchos años. Concedo que si uno ya ha llegado a la
conclusión de que la vida debe haber sido creada de manera
sobrenatural, no va a demostrar mucho entusiasmo en la
prosecución de mucho más trabajo experimental acerca del
tema. Pero si uno adopta una actitud más agnóstica,
entonces podría interesarse en llevar a cabo experimentos
adicionales.
Más
tarde o más temprano surgiría la cuestión de quién
o qué creó la vida. ¿Está la
inteligencia creadora totalmente dentro del cosmos, o es ella
(Él) el mismo que el Dios trascendente de la Biblia? Y luego
tenemos la cuestión de cómo aquella agencia
inteligente creó la vida (o incluso el mismo cosmos). Pero desde
luego los detalles de cómo una inteligencia sobrenatural
creó la realmente la vida no son susceptibles de
indagación por medio del método científico, y esto
es anatema para muchos científicos. Comentando acerca de la
evidencia de un propósito inteligente inmanente en el universo,
y la desgana de muchos científicos a aceptar tal perspectiva,
Hoyle (1983, pág. 9) escribía:
Esto es precisamente lo que los científicos ortodoxos
no están dispuestos a admitir. Porque podría resultar que
hubiera —a falta de una palabra mejor— connotaciones religiosas, y
porque los científicos ortodoxos están más
interesados en impedir un regreso a los excesos religiosos del pasado
que en buscar la verdad, la perspectiva nihilista descrita
previamente ha dominado el pensamiento científico a lo largo del
siglo pasado. (Énfasis añadido.)
Los científicos ortodoxos están
más interesados en impedir un regreso a los excesos religiosos
del pasado que en buscar la verdad.
Quizá
tendremos que afrontar a su debido tiempo la intensa posibilidad de que
el problema del origen de la vida revela una limitación del
mismo método científico. Si el método
científico no puede resolver todos los problemas (y,
¿quién mantiene que puede resolverlos todos?),
¿por qué entonces no podría ser que la gran
cuestión de los detalles físicos del origen de la vida
sea uno de estos problemas que está más allá del
alcance de nuestra metodología? Y en realidad,
¿cómo podemos eliminar esta posibilidad, dada la actual
situación en los estudios acerca del origen de la vida? Antes
que reconocer reconocer este tipo de limitaciones, muchos
preferirían seguir esforzándose, tejiendo una creciente
red de improbabilidades relacionadas, de énfasis mal situados, y
de extrapolaciones injustificadas, sólo para crear la
ilusión de que no hay problemas ahí. Pero bien al
contrario hay muchas dificultades en esta área de
investigación, y ya es hora que las afrontemos, tal como lo han
hecho Thaxton y sus colegas de manera valerosa. Pero las posibles
consecuencias no son insignificantes, y creo que esto tiene que ser
reconocido a cierto nivel por parte de los que ahora están
llevando a cabo investigaciones acerca del origen de la vida.
Notas
1. Un proceso «naturalista» es aquel que no
precisa de «ayuda» desde una fuente sobrenatural, sino que
tiene lugar de manera espontánea en virtud de las propiedades
químicas y físicas de la materia y energía en el
sistema. Vuelve al texto
2. Siguiendo este enfoque, yo hubiera podido llegar a la
conclusión de que el O2
estaba presente en la atmósfera primordial debido a que
se precisa de O2 para la
formación de microesferas de aldocianoina (Kenyon y Nissenbaum,
1976; Pollack y Heiderer, 1979). Vuelve al texto
3. Esto es especialmente evidente en el caso del origen
de las preferencias de isómeros ópticos. Las resoluciones
ópticas eficaces en laboratorio exigen, todas ellas, una
interferencia del investigador impermisible en experimentos de
simulación prebiótica (Wilder-Smith, 1981, págs.
15-23). Vuelve al texto
4. En recientes versiones del experimento de
simulación tipo Miller se omite de todas maneras el H2 en muchas
ocasiones por razones de tipo teórico. Debo mi reconocimiento al
Prof. Thwaites por señalarme aquí el peligro potencial. Vuelve al texto
Referencias
Ambrose,
E.J. (1982). The Nature and Origin of the Biological World.
Halsted Press, New York.
Bonner,
W.A. (1972). Origins of Molecular Chirality. En: Exobiology, C.
Ponnamperuma, ed., págs. 170-234, North Holland, Amsterdam.
Bonner,
W.A., Blair, N.E. y Lemmon, R.M. (1980). The Radioracemization of Amino
Acids by Ionizing Radiation: Geochemical and Cosmochemical
Implications. En: Limits of Life: Proceedings of the Fourth College
Park Colloquium on Chemical Evolution, C. Ponnamperuma y L.
Margullis, eds., págs. 183-194, D. Reidel, Dordrecht.
Cairns-Smith,
A.G. (1982). Genetic Takeover and The Mineral Origins of Life.
Cambridge University Press, Cambridge.
Clemmey,
H. y Badham, N. (1982). Oxygen in the Precambrian Atmosphere: An
Evaluation of the Geological Evidence. Geology, 10, 141-146.
Corliss,
J.B., Baross, J.A. y Hoffman, S.E. (1981). A Hypothesis Concerning the
Relationship Between Submarine Hot Springs and the Origin of Life on
Earth. Oceanologia Acta, SP, 59-67.
Crick,
F. (19810. Life Itself: Its Nature and Origin. Simon and
Schuster, New York.
Dikerson,
R.E. (1978). Chemical Evolution and the Origin of Life, Scientific
American, 231, n(o) 9, 70-86.
Dimroth,
E. y Kimberley, M.M. (1976). Precambrian Atmospheric Oxygen: Evidence
in the Sedimentary Distributions of Carbon, Sulphur, Uranium and Iron. Canadian
Journal of Earth Sciences, 13, 1161-1185.
Eden,
M. (1967). Inadequacies of Neo-Darwinian Evolution as a Scientific
Theory. En: Mathematical Challenges to the Neo-Darwinian
Interpretation of Evolution, P.S. Moorhead y M.M. Kaplan, eds.,
págs. 5-12, Wistar Institute Press, Philadelphia.
Eigen,
M. (1971). Selforganization of Matter and the Evolution of Biological
Macromolecules. Naturwiss. 58, 465-523.
Eigen,
M., Gardiner, W., Schuster, P. y Winkler-Oswatitsch, R. (1981). The
Origin of Genetic Information. Scientific American, 244, n(o)
4, 78-94.
Ellis,
G.P. (1959). The Maillard Reaction, En: Advances in Carbohydrate
Chemistry, M.L. Wolfram, ed., Vol. XIV, págs. 63-134.
Academic Press, New York.
Folsome,
C.E. (1979). The Origin of Life: A Warm Little Pond. Freeman,
San Francisco.
Folsome,
C.E., Allen, R.D., y Ichinose, N. (1975). Organic Microstructures as
Products of the Miller-Urey Electric Discharges. Precam. Res.,
2, 263-275.
Fox,
S. W. (1956). Evolution of Protein Molecules and Thermal Synthesis of
Biochemical Substances. American Scientist, 44, 337-347.
Fox,
S.W. (1964). Thermal Polymerization of Amino Acids and Production of
Formed Microparticles on Lava. Nature, 201, 336-337.
Fox,
S.W. (1974). The Proteinoid Theory of the Origin of Life and Competing
Ideas. American Biology Teacher, 36, 161-181.
Fox,
S.W. (1978). The Origin and Nature of Protolife. En: The Nature of
Life, W.H. Heidcamp, ed., págs. 23-92, University Park
Press, Baltimore.
Fox,
S.W. (1981). Origins of the Protein Synthesis Cycle. Int. J.
Quantum Chem. QBS, 8, 441-454.
Fox
S.W. y Dose, K (1977). Molecular Evolution and the Origin of Life,
2(a) Ed., Marcel Dekker, New York.
Fox,
S.W., y Harada K. (1960). The Thermal Copolymerization of Amino Acids
Common to Protein. J. Am. Chem. Soc., 82., 3745-3751.
Fox,
S.W., y Nakashima, T. (1980). The Assembly and Properties of
Protobiological Structures: The Beginning of Cellular Peptide
Synthesis. BioSystems, 12, 155-166.
Fox,
S.W., y Yuyama, S. (1964). Dynamic Phenomena in Microspheres from
Thermal Proteinoid. Comp. Biochem. Physiol., 11, 317-321.
Hoyle,
F. (1983). The Intelligent Universe: A New View of Creation and
Evolution. Holt, Rinehart and Winston, New York.
Hoyle,
F., y Wickramasinghe, N.C. (1981). Evolution from Space. J.M.
Dent, Londres.
Kenyon,
D.H. (1984a). A Comparison of Proteinoid and Aldocyanoin Microsystems
as Models of the Primordial Protocell. En: Molecular Evolution and
Protobiology, K. Matsuno, K. DOse, K. Harada y D.L. Rohlfing, eds.,
págs. 163-188. Plenum Press, New York.
Kenyon,
D.H. y Nissenbaum, A. (1976). Melanoidin and Aldocyanoin Microspheres:
Implications for Chemical Evolution and Early Precambrian
Micropaleontology, J. Mol. Evol., 7, 245-251.
Kenyon,
D.H. y Steinman, G. (1969). Biochemical Predestination,
McGraw-Hill, New York.
Kuppers,
Bernd-Olaf (1983). Molecular Theory of Evolution: Outline of a
Physico-Chemical Theory of the Origin of Life. Springer-Verlag,
Berlín.
Matsuno,
K., Dose, K., Harada, K., y Rohlfing, D.L., eds., (1984). Molecular
Evolution and Protobiology. Plenum Press, New York.
Miller,
S.L. (1953). A Production of Amino Acids under Possible Primitive Earth
Conditions. Science, 117, 528-529.
Miller,
S.L. (1955). Production of Some Organic Compounds under Possible
Primitive Earth Conditions. J. Am. Chem. Soc. 77, 2351-2361.
Miller,
S.L. (1957). The Mechanism of Synthesis of Amino Acids by Electric
Discharges. Biochem. Biophys. Acta, 23, 480-489.
Miller,
S.L. (1984). The Prebiotic Synthesis of Organic Molecules and Polymers.
En: Aspects of Chemical Evolution, C. Nicolis, ed.,
págs. 85-107, John Wiley and Sons, New York.
Miller,
S.L. y Orgel, L.E. (1974). The Origins of Life on the Earth.
Prentice-Hall, Englewood Cliffs, New Jersey.
Monod,
J. (1972). Chance and Necessity, Knopf, New York.
Nissenbaum,
A., Kenyon, D.H. y Oró, J. (1975). On the Possible Role of
Organic Melanoidin Polymers as Matrices for Prebiotic Activity. J.
Mol. Evol. 6, 153-170.
Parker,
g.E. (1980). Creation: The Facts of Life. CLP Publishers, San
Diego.
Pollock,
G.E. y Heiderer, R. (1979). Aldocyanoin Microspheres: Partial Amino
Acid Analysis of the Microparticulates Formed From Simple Reactants
Under Various Conditions. J. Mol. Evol., 13, 253-264.
Ring,
D., Wolman, Y., Friedmann, N. y Miller, S.L. (1972). Prebiotic
Synthesis of Hydrophobic and Protein Amino Acids. Proc. Nat. Acad.
Sci. U.S.A., 69, 765-768.
Sagan,
C. (1980). Cosmos. Random House.
Shapiro,
R. (1984). The Improbability of Prebiotic Nucleic Acid Synthesis. Origins
of Life, 14, 565-570.
Thaxton,
C.B., Bradley, W.L., y Olsen, R.L. (1984). The Mystery of Life's
Origin: reassessing Current Theories. Philosophical Library, New
York.
Wilder-Smith,
A.E. (1970). The Creation of Life: A Cybernetic Approach to
Evolution. Harold Shaw, Wheaton.
Wilder-Smith,
A.E. (1981). The Natural Sciences Know Nothing of Evolution.
Master Books, San Diego.
Yockey,
H. (1977). A Calculation of the Probability of Spontaneuos Biogenesis
by Information Theory. J. Theoret. Biol. 67, 377-398.
Yockey,
H. (1981). Self-Organization Origin of Life Scenarios and Information
Theory. J. Theoret. Biol. 91, 13-31.
 Arriba un nivel Arriba un nivel
De vuelta al índice general
De vuelta a la página principal
©
SEDIN 1997
Autor:
Dean H. Kenyon
Departamento de Ciencias Biológicas
Universidad Estatal de San Francisco
San Francisco, California 94132
FUENTE: ORIGINS
RESEARCH, Vol. 12 - Número 1, Primavera/Verano 1989
Traducción del inglés: Santiago Escuain
© Santiago Escuain 1997, por la traducción
© Copyright SEDIN 1997 para el formato electrónico -
www.sedin.org. Este texto se puede reproducir libremente para fines no
comerciales y citando la procedencia y dirección de SEDIN,
así como esta nota en su integridad.
SEDIN
Servicio Evangélico - Documentación - Información
Apartat 2002
08200 SABADELL
(Barcelona) ESPAÑA
|
Lecturas complementarias
David Coppedge
El
naufragio en las investigaciones sobre el origen de la vida (2007)
Nancy Pearcey
El
primer paso
La Evolución Química -
Entrevista con el doctor Charles B. Thaxton - Coautor de The Mystery of
Life’s Origin
(1985)
William
E. James, Ph. D.
Santiago Escuain
El Origen del Primer Sistema Vivo (1977-1984)
Duane T. Gish, Ph.D.
Especulaciones y
experimentos sobre el Origen de la vida: Crítica (1972)
Visite también:
El Museo de Máquinas Moleculares
Índice de
boletines
Índice de línea
sobre línea
Página principal
Índice general
castellano
Libros recomendados
orígenes
vida cristiana
bibliografía general
Coordinadora Creacionista
Museo de Máquinas
Moleculares
Temas de actualidad
Documentos en PDF
(clasificados por temas)




|