Tomado y traducido de ARN - Access Research Network [Véase al pie]
EL FLAGELO BACTERIANO
Un ejemplo de complejidad irreducible
Para entrar en el
Vestíbulo del Museo de Máquinas Moleculares, haga clic aquí.
Para ver una animación del flagelo
bacteriano, haga clic aquí.
El
flagelo bacteriano es un ejemplo de lo que Michael Behe describe como
un sistema irreduciblemente complejo. En su libro La Caja Negra de Darwin explica él que
estos sistemas de complejidad irreducible no podrían haber
surgido mediante un proceso darwiniano de pasos graduales.
Debido a que el flagelo bacteriano está
necesariamente compuesto de al menos tres piezas --una paleta, un rotor
y un motor-- es irreduciblemente complejo. La evolución gradual
del flagelo, como la del cilio, se enfrenta por ello a unos
obstáculos gigantescos. (p. 72)
Behe
sumariza la estructura del flagelo bacteriano en estos términos:
Algunas bacterias presentan un maravilloso dispositivo para
la natación, el flagelo, que no tiene semejante en
células más complejas. En 1973 se descubrió que
algunas bacterias nadan haciendo girar sus flagelos. De modo que el
flagelo bacteriano actúa como un propulsor giratorio, en
contraste con el cilio, que actúa más a semejanza de un
remo.
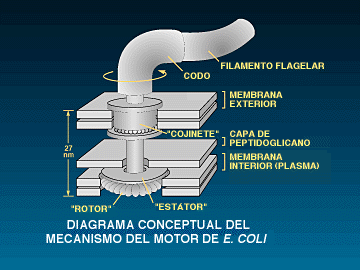
La estructura de un flagelo es muy diferente de la de un
cilio. El flagelo es un largo filamento semejante a un cabello
empotrado en la membrana de la célula. El filamento externo se
compone de un solo tipo de proteína, llamada
«flagelina». El filamento de flagelina es la superficie de
la paleta que contacta con el líquido durante la
natación. Al final del filamento de flagelina, cerca de la
superficie de la célula, hay una protuberancia en el grosor del
flagelo. Es aquí que el filamento se une a la transmisión
del rotor. La unión material está compuesta de una pieza
llamada «proteína de codo», El filamento del flagelo
bacteriano, a diferencia del cilio, no contiene proteína motora;
si se arranca, el filamento flota rígido en el agua. Por ello,
el motor que hace girar el filamento propulsor tiene que estar situado
en algún otro lugar. Mediante experimentos se ha comprobado que
está situado en la base del flagelo, donde la microscopía
electrónica revela la presencia de varias estructuras en forma
de anillo. La naturaleza rotatoria del flagelo tiene unas claras e
ineludibles consecuencias ... (pp. 70-72)
Las
consecuencias a que se refiere Behe se deducen de la naturaleza de sus
componentes, que presentan una complejidad irreducible, y cuyo
descubrimiento socava cualquier explicación darwinista de los
orígenes. Behe llega a esta conclusión:
Sumarizando, al comenzar los bioquímicos a examinar
unas estructuras aparentemente simples como los cilios y los flagelos,
han descubierto una abrumadura complejidad, con docenas e incluso
cientos de piezas diseñadas con precisión. Es muy
probable que muchas de estas piezas que no hemos considerado
aquí sean todas ellas imprescindibles para que cualquier cilio
funcione en una célula. Al aumentar el número de piezas
necesarias, la dificultad de erigir el sistema de manera gradual
aumenta exponencialmente, y la probabilidad de escenarios indirectos se
desploma. Darwin parece más y más impotente. La nueva
investigación acerca de los papeles de las proteínas
auxiliares no puede simplificar el sistema complejo irreducible. No se
puede aminorar la intransigencia del problema, sólo hará
que empeorar. La teoría darwinista no ha dado explicación
al cilio ni al flagelo. Y la abrumadora complejidad de los sistemas de
natación nos impulsa a pensar que nunca podrá darle una
explicación. (p. 73)
Behe
concluye que estos sistemas irreduciblemente complejos son en
último término resultado de un diseño inteligente.
(Se
debería observar que Behe no tiene objeciones al concepto de un
ancestro común universal. Sus objeciones a la evolución
se limitan al rechazo del mecanismo neodarwinista como
explicación suficiente para el origen de todos los sistemas
biológicos.)
Objeciones a la tesis del diseño inteligente
La
mayoría de los científicos argumentarán que el
«diseño inteligente» no es una explicación
puramente naturalista o materialista del origen de la
información biológica y que por ello mismo no es una
explicación científica válida. Escondido a menudo
en este argumento encontramos la presuposición filosófica
o teológica de que Dios (o cualquier otro agente inteligente) no
tuvo papel alguno activo, directo o detectable en la creación de
ningún sistema biológico. Esta presuposición exige
que toda la información genética tiene que haber surgido
sobre una base totalmente al lazar y/ gradual, paso a paso. De ello
seguiría que la naturaleza ha de ser un continuo y que la
historia natural ha de estar representada por un árbol de la
vida. Encerrada en el naturalismo, la hipótesis de una
descendencia común tiene que ser aceptada de manera
axiomática y por ello no está sujeta a la
falsación científica.
Cosa
irónica, los científicos quedan más limitados bajo
un paradigma naturalista que bajo un paradigma de diseño
inteligente. El diseño inteligente del universo pudiera haber
incluido la creación de leyes naturales y de condiciones
iniciales suficientes para permitir la evolución de un solo
árbol de vida sobre la tierra. Pero el diseño inteligente
pudiera también haber incluido puntuaciones a lo largo de la
historia natural, actos directos de creación o de
infusión de nueva información genética, que
resultaría en discontinuidades naturales. Ejemplos de estas
discontinuidades naturales podrían incluir el origen de la vida,
el origen de las categorías taxonómicas más
elevadas (esto es, el origen de la mayoría de los fílums
en la explosión cámbrica de vida), o el origen de los
sistemas irreduciblemente complejos. Si existen discontinuidades
naturales, la historia natural quedaría modelada con más
precisión como un bosque de la vida. Lo importante es que un
paradigma de diseño inteligente puede acomodar tanto un
árbol como un bosque de la vida así como la existencia de
procesos que o bien produzcan o bien limiten el cambio evolutivo. Pero
a diferencia del paradigma naturalista, el diseño inteligente
demanda que los científicos desarrollen hipótesis
susceptibles de prueba para determinar cuál es el modelo que
mejor se ajusta a los datos.
La
meta de la ciencia es la descripción y modelado más
precisos posibles del Cosmos. En tanto que puede que haya evidencias
circunstanciales que apoyen bien un modelo continuo bien un modelo
discontinuo de la vida, es de vital importancia que reconozcamos
cómo las presuposiciones de partida afectan las conclusiones a
las que uno llega. Desde una presuposición inicial de
diseño inteligente o de agnosticismo filosófico (ambas
permiten que el Cosmos sean bien continuo, bien discontinuo) el
científico tiene evidentemente una mayor libertad de pensamiento
que la que se ofrece desde un paradigma puramente naturalista. Junto
con esta libertad hay la responsabilidad de poner a prueba lo que los
filósofos naturalistas simplemente dan como supuesto.
¿Actúa la Selección Natural
inhibiendo los grandes cambios evolutivos?
Hay
numerosos ejemplos de organismos con sistemas de componentes
interdependientes y muy específicos, todos los cuales han de
quedar integrados antes que sean funcionales y ofrezcan cualquier
ventaja selectiva. La naturaleza está repleta de estos sistemas
de «complejidad irreducible». Aunque pueda haber algunos
ejemplos en los que ciertos componentes puedan presentar alguna
función preadaptativa, estos casos pueden considerarse como
excepciones a una regla más general. Como norma, cualquier
subconjunto de los componentes resultaría ser una carga para el
organismo donde ocurriera, y sería por ello eliminado por la
selección natural. Tomemos por ejemplo el mecanismo sensorial y
motor de la bacteria común Escherichia coli, un organismo
unicelular procariota relativamente simple.
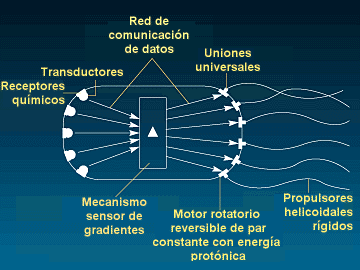
Esquema conceptual de los sistemas sensoriales y de
navegación de la Escherichia coli
El
mecanismo sensorial y motor se compone de un número de
receptores que detecta inicialmente las concentraciones de una
diversidad de sustancias químicas. Unos componentes secundarios
extraen información de estos detectores que a su vez se emplea
como entrada a un mecanismo detector de gradientes. La salida de este
mecanismo se emplea para impulsar un conjunto de motores rotatorios
reversibles de par constante de energía protónica que
transfieren su energía mediante un tren de transmisión
microscópico y que hacen girar unos flagelos helicoidales. Este
sistema tan sumamente integrado permite que la bacteria se desplace a
una velocidad de aproximadamente diez longitudes corporales por
segundo. El doctor Robert Macnab de la Universidad de Yale
concluía un estudio de 50 páginas acerca de este
mecanismo con estas observaciones:
Como comentario final, no podemos más que
maravillarnos ante la complejidad, en una sencilla bacteria, de todo el
sistema sensorial y motor que ha sido el tema de esta reseña, y
hacer la observación de que nuestro concepto de evolución
por ventajas selectivas es evidentemente una enorme
simplificación. Por ejemplo, ¿qué ventaja
podría haber en un «pre-flagelo» (o sea, un
subconjunto de sus componentes)?, y, con todo, ¿cuál
sería la probabilidad de un desarrollo
«simultáneo» del orgánulo a un nivel en el
que devenga ventajoso?
Macnab, R. (1978), «Bacterial Mobility and
Chemotaxis: The Biology of a Behavioral System [Movilidad y quimiotaxia
bacteriana: La biología de un sistema de conducta]»,
CRC Critical Reviews in Biochemistry, 5, 1978, págs. 291-341.
Lecturas adicionales
Una
excelente introducción general a los flagelos puede encontrarse
en Voet, D. and Voet, J. G. (1995) Biochemistry, 2nd edition,
John Wiley and Sons, New York, pp. 1259-1260.
En
las siguientes referencias se pueden encontrar mayores detalles acerca
del motor flagelar:
Schuster,
S. C. and Khan, S. (1994) "The Bacterial Flagellar Motor," Annual
Review of Biophysics and Biomolecular Structure, 23, 509-539;
Caplan, S. R. and Kara-Ivanov, M. (1993) "The Bacterial Flagellar
Motor," International Review of Cytology, 147, 97-164.
Para
un análisis de las pretensiones de Nicholas Matzke y Kenneth
Miller de haber dado respuesta a las tesis de Michael Behe,
véase:
Traducción al castellano: Santiago Escuain
Copyright de la traducción © 2001 SEDIN. Todos los derechos
reservados.
Este fichero se puede reproducir en su integridad para
usos no comerciales.
Se agradecerán enlaces con los sitios Web de Access Research
Network y SEDIN.
Artículo original en inglés: Pulse
aquí
Page development by Premier Publications
for Access Research Network,
Colorado Springs, Colorado, USA.
[ Museo de Máquinas Moleculares ] [ Página anterior ] [ Índice general ] [ Vuelve
a la página principal ]
[ Previous Page ] [ Molecular Machines Museum (En
inglés) ] [ Página Principal de ARN
[Access Research Network] ]
SEDIN
Servicio Evangélico - Documentación - Información
Apartado 2002
08200 SABADELL
(Barcelona) ESPAÑA
www.sedin.org
|
Índice:
Índice de boletines
Índice de línea
sobre línea
Página principal
Índice general castellano
Libros recomendados
orígenes
vida
cristiana
bibliografía
general
Coordinadora Creacionista
Museo de Máquinas Moleculares
Temas de actualidad
Documentos en PDF
(clasificados por temas)




|