|
Servicio
Evangélico de Documentación e
Información
línea sobre línea |
|||||||||| Apartado 2002 - 08200 SABADELL (Barcelona) ESPAÑA | SPAIN ||||||||
Duane T. Gish, Ph.D.
|
|
MODELO «CREACIÓN» |
MODELO «EVOLUCIÓN» |
|
Por Actos de un Creador |
Por procesos naturales mecanicistas
debidos a propiedades inherentes a la materia inanimada. |
|
Creación de géneros básicos de vegetales
y animales con características ordinales completas en los primeros
representantes. |
Origen de todo lo viviente a partir de una sola fuente de vida que surgió por si misma de la materia inanimada. |
|
Variación y especiación limitada dentro de cada género. |
Variación ilimitada. Todas las formas están relacionadas genéticamente. |
Estos dos modelos nos permitirían, a su vez, emitir las siguientes predicciones concernientes al registro fósil:
|
MODELO «CREACIÓN» |
MODELO «EVOLUCIÓN» |
|
Aparición repentina de una gran variedad de formas altamente complejas. |
Cambio gradual de las formas más simples
a formas más y más complejas. |
|
Aparición repentina de todos los géneros creados con características ordinales completas. Discontinuidades bruscas separando grupos taxonómicos principales, sin formas transicionales entre categorías principales de transición |
Series transicionales conectando todas las categorías. No debe haber discontinuidades sistemáticas. |
COMPARACIÓN DEL REGISTRO FÓSIL CON LAS PREDICCIONES DE LOS DOS MODELOS
1. Del Precámbrico al Cámbrico:
Las rocas más
antiguas en las cuales se encuentran fósiles indiscutibles son las del período
Cámbrico. En estos depósitos sedimentarios se encuentran miles y miles de
millones de fósiles de formas altamente complejas de vida. Esta incluyen
esponjas, corales, calamares, gusanos, moluscos, crustáceos. De hecho. se han
encontrado todas las clasificaciones principales de formas invertebradas de vida
en las rocas Cámbricas. Estos animales son tan altamente complejos que se estima
que precisaron de 1.500 millones de años para evolucionar.
Y, ¿qué es lo que en contramos en las rocas Precámbricas, de mayor antigüedad que las Cámbricas? ¡No se ha encontrado ni un solo fósil indiscutible en ellas! Se puede decir con toda certeza que los antepasados de la fauna Cámbrica, si han existido alguna vez, jamás han sido hallados. [23] , [24] , [25]
A pesar de las afirmaciones en contra, los reportajes concerniendo el descubrimiento de microfósiles (bacterias y algas unicelulares microscópicas) en rocas Precámbricas, a las que se las ha asignado una edad de 1.000 a 2.000 millones más de años que a las del Cámbrico, son cuestionables, y desde luego están abiertos a discusión. Algunos artículos recientes tienden a exponer las incertidumbres involucradas en tales identificaciones. [26] , [27] , [28] Por ejemplo, a pesar de que aceptan la probabilidad de que alguno de estos pretendidos microfósiles de la era Precámbrica sea de origen biológico, Engel, et al, han advertido que:
Aun si estos pretendidos microfósiles representan restos de formas genuinamente Precámbricas, nos encontramos de todas maneras con una discontinuidad tremenda entre las formas unicelulares microscópicas de vida, y las muy diversificadas y altamente complejas formas de vida encontradas en el Cámbrico, una discontinuidad que se extiende supuestamente sobre mil o dos mil millones de años en las eras geológicas.
Axelrod
afirma, refiriéndose a este problema:
«Uno de los principales problemas no resueltos de Geología y Evolución es la presencia de invertebrados marinos multicelulares diversificados en rocas Cámbricas Anteriores en todos los continentes, y su ausencia en rocas de mayor antigüedad.»
Después de
considerar los varios tipos que se hallan en el Cámbrico, Axelrod
prosigue:
«No obstante,
cuando vamos a las rocas Precámbricas para hallar los antecesores de estos
fósiles Cámbricos, no se encuentran por ninguna parte. Se conoce, ahora,
que muchas secciones de rocas sedimentarias (de unos 1.600 m.) descansan en
sucesión ininterrumpida bajo estratos que contienen los más primitivos fósiles
Cámbricos. Aparentemente, estos sedimentos fueron adecuados para la preservación
de fósiles, a causa de que a menudo son idénticas a los de las formaciones
fosilíferas que descansan encima de ellos y, a pesar de ello, no se
encuentran fósiles en ellos».25
Según todas las apariencias, basados en los hechos conocidos del registro fósil, hubo una repentina explosión de vida con un alto nivel de complejidad. El registro fósil no nos da ninguna evidencia de que estos animales Cámbricos se desarrollaran a partir de precedentes formas ancestrales. Aun más, no se ha encontrado ningún fósil que pueda ser considerado como forma de transición entre los grupos principales de clasificación que lo forman. En su aparición más primitiva, estas categorías de vida invertebrada eran ya tan «divergentes» con tanta claridad como hoy en día.
¿Cómo queda el modelo evolucionista a la luz de estos hechos? Obviamente, están en completa contradicción con sus afirmaciones. Esto lo ha admitido el mismo George Gaylord Simpson, que declaró:
Simpson ha
luchado valientemente, pero no fructuosamente, con este problema, habiéndose
visto obligado a conceder que la ausencia de fósiles Precámbricos (aparte de los
pretendidos microfósiles) es
No obstante, estos hechos están totalmente de acuerdo con, el modelo «creación». El registro fósil revela una aparición repentina de gran variedad de formas altamente especializadas y complejas, sin antecesores evolutivos y sin formas de transición que conecten los grupos taxonómicos mayores, tal como queda postulado en el modelo creacionista, pero contradicen incuestionablemente las predicciones del modelo evolucionista.
El resto de la historia de la vida revela una ausencia destacable de las muchas formas de transición exigidas por la teoría. De hecho, hay una deficiencia sistemática de formas de transición entre las categorías mayores de clasificación, tal como está predicho en el modelo creacionista.
2. De los
Invertebrados a los Vertebrados:
La idea de que
los vertebrados derivaron de los invertebrados es una pura conjetura que no
puede ser documentada por el registro fósil. Se ha propuesto, en base de la
anatomía comparativa y de la embriología de las formas vivientes, casi cada uno
de los grupos invertebrados en ocasiones diferentes. [31] , [32]
Supuestamente, la forma de transición de invertebrado a vertebrado
pasó por una etapa cordada sencilla.
¿Posee el registro fósil de evidencias de tal transición? La respuesta a esta pregunta tiene que ser un NO absoluto.
Ommaney ha afirmado:
¡Increíble!
¡Cien millones de años de evolución, y ninguna forma de transición! Todas las
hipótesis combinadas, no importa lo ingeniosas que sean, nunca pueden pretender,
basándose en, la teoría de la evolución, explicar una discontinuidad de tal
magnitud. Por otra parte, están de completo acuerdo con el modelo
creacionista.
3. El problema del origen de las diversas clases de Peces:
Una lectura cuidadosa del libro de Romer, Paleontología Vertebrada, 30 parece no permitir otra conclusión sino la de que las clasificaciones principales de peces son ya clara y distintamente «divergentes», sin formas de transición que las conecten. La primera en aparecer en el registro fósil es la clase Agnatha. Los más antiguos de estos vertebrados, representantes de los dos órdenes Osteostraci y Heterostraci, han quedado casi siempre incrustados en material óseo u otro material duro. En lo que concierne a su origen, Romer escribe:
«En los sedimentos de la época Silúrica posterior y Devónica anterior están presentes muchos vertebrados de características ictíneas de varios tipos, y es obvio que ha tenido que existir una larga historia evolutiva antes de este tiempo. Pero en lo que se trata de esta historia, la ignoramos mayormente» (p. 15, edición inglesa).
En cuanto a los Ostracodermos (Osteostraci), Romer escribe así:
«Cuando primeramente aparecen estos Ostrocodermos, ya tienen una larga historia a sus espaldas y están divididos en varios grupos distintos» (p. 16).
De los Heterostraci, dice Romer que desde luego no están relacionados de cerca con las otras formas de la clase Agnatha. Si evolucionaron, también deben tener una historia evolutiva muy larga. Pero, igual que los Ostracodermos, aparecen repentinamente en el registro fósil sin ninguna evidencia de antecesores evolutivos.
Los Placodermos son causantes de buenos dolores de cabeza. Existían dentro de los Placodermos sobre unas seis clasificaciones mayores de peces monstruo. De ellos dice Romer:
Después de esto, sigue diciendo:
«Aparecen en un momento –más o menos sobre la frontera Siluriano-Devónica– en que esperaríamos la aparición de los antecesores apropiados de los tiburones y de los grupos superiores de peces óseos. Serían de esperar formas «generalizadas» que encajaran limpiamente en nuestra preconcebida secuencia evolutiva. ¿Las conseguimos en los Placodermos? No, en absoluto. En su lugar encontramos una serie de tipos brutalmente imposibles que no encajan en ningún esquema apropiado, que no parecen venir de ninguna fuente anterior –a primera vista–, ni ser antecesores apro piados para ningún tipo posterior más avanzado. De hecho, lo que uno tiende a sentir es que la presencia de estos Placodermos, que forman una parte tan importante de la historia Devónica, es un episodio incongruente. ¡La situación se habría simplificado si nunca hubieran existido!» (p. 33).
Pero existieron, y su registro no apoya, sino que contradice fuertemente él modelo evolucionista.
Los tipos superiores de peces, los «ortodoxos», estructurados bajo planes bien reconocidos, con aletas apareadas y mandíbulas bien desarrolladas, están colocados dentro de las dos clases, los Chondrichthyes o peces cartilaginosos, y los Osteichthyes, o peces óseos superiores.
Algunos, en el pasado, han argumentado que la ausencia de huesos en los peces cartilaginosos representa una condición primitiva, y que los Chondrichthyes fueron una etapa evolutiva precediendo a los peces óseos. Romer argumenta con fuerza contra esta línea de pensamiento, señalando el hecho de que los tiburones son unos de los últimos grupos de clasificación taxonómica mayores en aparecer en el registro fósil.
Así, Romer continúa:
«El registro, de hecho, encaja mejor en la conjetura opuesta: que los tiburones son degenerados en lugar de primitivos en sus caracteres esqueléticos; que su evolución ha sido paralela a la de otros varios tipos de peces en su tendencia hacia la reducción ósea; y que se tiene que buscar a sus antecesores entre los peces óseos primitivos con mandíbulas, del tipo general de los Placodermos. Ningún Placodermo conocido puede ser identificado como antecesor verdadero de los Chondrichthyes, pero nos hemos dado cuenta de que algunos de los peculiares Petalichthydeos parecen mostrar etapas morfológicas de reducción esqueletal. El conocimiento creciente de los Placodermos Devónicos anteriores puede algún día atar cabos» (p. 38).
Antes, y refiriéndose a los Placodermos, Romer había declarado:
«Debemos considerar seriamente que al menos los Tiburones y Quimeras puedan haber descendido de tales imposibles antepasados» (p. 34). Una consideración al modelo Creacionista, ¿no sería aquí mucho más razonable que una apelación a imposibles antepasados?
Sobre los peces típicamente óseos, Romer registra el hecho de que su aparición en el registro fósil es «dramáticamente repentina» (p. 52). Más adelante (p. 53), afirma:
«El antecesor común de los grupos de peces óseos es desconocido.» «Hay varios caracteres, muchos de ellos descritos anteriormente, en los cuales las dos típicas subclases de peces óseos ya son claramente divergentes cuando las contemplamos en su principio.»
Así, el registro fósil no muestra antecesores ni formas de transición de los grupos mayores de clasificación de peces. Basándonos en el registro conocido, tales antecesores hipotéticos y las formas de transición necesarias son tan sólo el producto de la especulación. ¿Cómo pues se puede presentar el argumento de que la explicación ofrecida por el modelo evolucionista es más científica que la ofrecida por el modelo creacionista?
Y, ¿no está de
acuerdo esta evidencia con el modelo creacionista, siendo a la vez un embarazo
para el modelo evolucionista?
4. De los Peces a los Anfibios:
Se ha investigado cuidadosamente el registro fósil, buscando series de transición que conectasen los peces con los anfibios, pero no se han hallado. La conexión más «próxima» que se ha propuesto ha sido la que pretendidamente existe entre el Crosopterigio Ripidistio (pez) y los anfibios del género Ichthyostega. Existe, no, obstante, una tremenda discontinuidad entre los Crosopterigios y los Ichthyostegas, una discontinuidad que se habría extendido durante muchos millones de años, y durante los cuales hubieran debido existir innumerables formas de transición. Estas formas de transición deberían mostrar un cambio lento y gradual de las aletas pélvicas y pectorales del Crosopterigio, transformándose en los pies y piernas del anfibio, juntamente con la pérdida de otras aletas, y la concurrencia de otras transformaciones requeridas para su adaptación al habitat terrestre.
¿Cuáles son los hechos? No se ha encontrado ni una forma de transición entre la aleta del Crosopterigio y el pie del Ichthyostega. Los miembros del Ichthyostega eran ya del tipo anfibio básico, no mostrando ningún vestigio de descendencia de aletas.
¿Por qué se eligió el Crosopterigio Ripidistio como el supuesto antepasado de los anfibios? Ante todo, no hay ninguno mejor a mano. Echando a faltar un candidato a intermediario entre peces y anfibios, se investigaron los varios grupos de peces. Entonces se adoptó el Crosopterigio como el grupo más aproximado antepasado de los anfibios. Esta elección se debió mayormente a unos diseños de cerebro similares al del Ichthyostega, a la posesión de la vértebra en «arco» encontrada en el Ichthyostega y en otros Labirintodontos, y la presencia de huesos en las aletas, por lo cual, en algunos aspectos, se podría construir una ruda homología con los huesos del cuadrúpedo.

Reconstrucciones de un anfibio Ichthyostega y su supuesto antepasado Crosopterigio.
Según Romer,
la presión selectiva que permitió el origen de los anfibios cuadrúpedos a partir
de sus antecesores los peces fue e1 hecho de que en el período Devónico fueron
características unas sequías periódicas, durante las cuales se supone que
evolucionaron los anfibios. Esto obligó al Crosopterigio a salir de los lagos y
ríos que se secaban (se cree que poseía pulmones) para buscar otros
parajes acuáticos. Aquellas formas que habían heredado cambios por mutaciones
que permitiesen una locomoción más eficiente por tierra habrían
sobrevivido en números mayores que formas menos bien equipadas. Un gran número
de episodios de estos dieron origen, después de millones de años, a un verdadero
anfibio. [34]
Esta historia, atractiva a primera vista, pierde su plausibilidad cuando se consideran los siguientes hechos: Ya que los anfibios se encuentran en el Devónico posterior, hubieran debido evolucionar bien dentro del Devónico cuando sus supuestos antepasados Crosopterigios estaban aún florecientes. Si la historia de Romer es cierta, en lugar de ello, él Devónico debería mostrar extinciones en masa de los Crosopterigios, así como de otras formas de agua dulce. Lo cierto es precisamente lo contrario.
En el Carbonífero Primitivo anterior se encuentran otros tres órdenes anfibios. Ya que estos anfibios altamente diversificados aparecen primeramente en el Carbonífero Primitivo anterior con sus características ordinales completas, tendrían que haber empezado a evolucionar bien atrás, en el Devónico, tal como supuestamente hicieron los Ichthyostégidos. Las mismas presiones selectivas debieron haber sido ejercidas sobre estos tres órdenes como sobre los Ichthyostégidos y deben haber evolucionado a partir de los Crosopterigios o de los Ichthyostégidos. Pero ninguno de estos tres órdenes tiene la vértebra en forma de «arco» que poseen los Crosopterígidos y los Ichthyostégidos, sino que poseen la vértebra «más primitiva», la del tipo «Lepospondilos» o vaina. ¿Cómo, pues, se puede utilizar la vértebra en «arco» para conectar el Crosopterigio con lo s anfibios?
Aún más, de estos tres órdenes, el Aistopoda poseía un cuerpo largo, como de serpiente, con unas 200 vértebras. En todas estas formas los pies eran pequeños, y muchas ni siquiera los poseían, ¡ni aun siquiera trazas de la cintura escapular o pélvica! En algunas formas del orden Nectridea los cuerpos eran también largos como los de los Aistópodos, faltando asimismo los pies. Si el Ichthyostega o una forma similar fue el antecesor de todos los anfibios, entonces tenemos que mientras que él estaba en el proceso de derivar cuatro pies a partir de las aletas pectorales y pélvicas de su antecesor Crosopterigio, ¡su descendencia estaba igual de ocupada intentando desembarazarse de ellos!
¿Qué presiones selectivas dieron lugar a los cuatro pies en los Ichthyostégidos mientras que causaban simultáneamente su reducción y pérdida entre los Aistópodos y Nectrídeos? ¿Por qué estas formas tan diversas aparecen en el registro fósil con una «divergencia» ya completa en su primera aparición, sin evidencia de formas de transición?
Los tres órdenes existentes de anfibios incluyen las salamandras y lagartijas (Urodelata o Caudata); los ápodos (Apoda o Caecilia), parecidos a los gusanos, sin órganos de locomoción; y las ranas y sapos (Anura o Salientia) que están entre los vertebrados terrestres, más especializados, con extremidades posteriores largas y sin cola. Todos estos modernos anfibios poseen la vértebra más «primitiva» del tipo Lepospondilos, en lugar de la. vértebra en «arco» que supuestamente conecta a los anfibios con sus, antecesores Crosopterigios. Aun más, no hay formas de transición conectando estos tres órdenes modernos que constituyen la subclase Lissamphibia, con los anfibios hallados en el Paleozoico. Refiriéndose a ellos, Romer afirma:
La discontinuidad extremadamente amplia entre pez y anfibio, como se ha observado entre los Crosopterigios Ripidistios y los Ichthyostégidos, la aparición repentina –de hecho– de todos los órdenes anfibios del Paleozoico con sus diversas características ordinales completas en los primeros representantes, la ausencia de cualquier forma de transición entre estos órdenes del Paleozoico, y la ausencia de formas de transición entre ellas y las que existen actualmente, es, todo ello, contradictorio a las predicciones del modelo evolucionista. No obstante, estos hechos son tal como los predichos por el modelo creacionista.
5. De los Anfibios a los Reptiles y Mamíferos:
Es en la frontera anfibiana-reptiliana y reptiliana-mamífera donde se han hecho las más fuertes reivindicaciones de tipos de transición que conecten clases. Pero al examinarlos con atención, vemos que son simplemente aquellos que, en el registro fósil, más se parecen en estructura esqueletal.
La conversión de un invertebrado a vertebrado, de pez a cuadrúpedo con pies y piernas, o de animal terrestre a volátil son unos pocos ejemplos de cambios que requerirían una revolución en toda la estructura del animal. Tales transformaciones deberían proveer una serie de transición claramente reconocible como tal en el registro fósil si hubieran ocurrido por medio de un proceso evolutivo. Por otra parte, si el modelo creacionista es el verdadero, es en estas fronteras donde la ausencia de formas de transición será más evidente.
Pero vemos que lo cierto es lo contrario en la frontera anfibio-reptiliana y en la reptiliano-mamífera, particularmente en la primera. Siendo posible distinguir entre los reptiles y anfibios actuales, vivientes, en base de sus características esqueletales, son –no obstante– mucho más distinguibles por medio de sus partes blandas y, de hecho, la característica definitiva que separa a los reptiles de los anfibios es la posesión por parte de los reptiles, en contraste con los anfibios, del huevo amniótico.
Muchas de las características para determinar un mamífero residen en su anatomía o fisiología «blanda». Esto incluye su manera de reproducirse, la temperatura caliente de la sangre, respiración por medio de la posesión de un diafragma, la posesión de glándulas mamarias y la posesión de pelo.
Con todo esto, las dos diferencias más fácilmente distinguibles entre reptiles y mamíferos no han sido nunca conectadas por series de transición. Todos los mamíferos, existentes o fósiles, tienen un solo hueso, el dentario, a cada lado de la mandíbula inferior, y todos los mamíferos, existentes o fósiles, tienen tres osículos auditivos (que forman el oído medio), el martillo, el yunque y el estribo. En algunos reptiles fósiles, el número y tamaño de los huesos de la mandíbula inferior es reducido en comparación con los reptiles vivientes. Aun así, todos los reptiles, vivientes o fósiles, tienen como mínimo cuatro huesos en la mandíbula inferior, y todos poseen un solo osículo auditivo, el estribo.
No hay, por ejemplo, formas de transición que muestren dos o tres huesos en la mandíbula, o dos osículos auditivos, y tampoco no ha explicado nadie como se las arreglaba la forma de transición para masticar mientras su mandíbula se tenía que dislocar y rearticular, o como podía oír mientras se le iban formando los otros dos huesos en el oído medio.
Aún hay más. Para que los hechos del registro fósil puedan encajar en las predicciones del modelo evolucionista se tiene que establecer una secuencia cierta de ti empo que encaje con estas predicciones. Esto no ha sido posible con la secuencia Anfibios-Reptiles-Mamíferos a base del material fósil descubierto hasta ahora.
Las conocidas formas Seymuria y Didactes, de las cuales se dice que están en la línea de división entre reptiles y anfibios, son del Pérmico anterior. O sea, que han llegado 20 millones de años demasiado tarde para ser los antecesores de los reptiles, si es que hemos de seguir la cronología evolucionista. Los pretendidos reptiles-base del orden Cotilosauria, se encuentran no en el Pérmico posterior a ellos, sino en el período precedente, el Carbonífero tardío (Pensilvánico).
De hecho, los reptiles «mamiferoides» del suborden Sinápsida, que supuestamente dieron origen a los mamíferos, se hallan en el Pensilvánico (Carbonífero tardío), y posiblemente en el Pensilvánico anterior. Así, el Seymuria y el Didactes, «antecesores» de los reptiles, no solamente postdatan a los mismos reptiles por decenas de millones de años, sino que también postdatan a los mismos antecesores de los mamíferos por un lapso de tiempo igual.
Según el modelo evolucionista, los mamíferos asumieron la supremacía sobre los reptiles en un período relativamente tardío de la historia de los reptiles. Si esto es cierto, se podría asumir razonablemente que la rama de reptiles de que tuvo su origen se desarrolló tardíamente en la historia de los reptiles. Pero aquí también vemos que lo cierto es lo contrario, si es que fueron los Sinápsidos los que antecedieron a los mamíferos. La subclase Sinápsida está fechada entre los más primitivos de los reptiles conocidos, no los últimos, y se supone que pasaron su apogeo aún antes de la aparición de los Dinosaurios.
Según Romer, los reptiles sinápsidos fueron disminuyendo en número durante el Triásico, quedando esencialmente extintos hacia el final de aquel período, y pasaron muchos millones de años antes de que sus «descendientes» llegaran a una posición de dominio. [36] Si la selección natural es el proceso que rige la evolución (y la definición de la Selección Natural es que es un proceso por el que el organismo más capacitado para la lucha por la vida es el que produce mayor descendencia), entonces, si la anterior historia de Romer es cierta, tendremos que concluir en que la transición del reptil al mamífero triunfó a pesar de la Selección Natural, en lugar de triunfar a causa de ella.
6) El Origen del Vuelo:
El origen del vuelo debería suministrarnos casos excelentes para presentar evidencias evolucionistas en contra de las tesis creacionistas, si lo primero fuera lo cierto. Para llegar a volar, casi todas las estructuras de un animal no volador requerirían un cambio de estructura, y las formas de transición resultant es deberían ser fácilmente detectables en el registro fósil. Se supone que el vuelo ha evolucionado separada e independientemente en cuatro ocasiones: en los insectos, aves, mamíferos (los quirópteros), y reptiles (los pterosaurios, ya extinguidos). En cada caso se supone que se precisaron muchos millones de años para llegar al vuelo, y en cada caso se hubieran visto envueltas en ello formas casi innumerables de transición. Pero aun así, no se encuentra nada, ni un solo caso, que se aproxime a una serie de transición.
E. C. Olson, un evolucionista y geólogo, afirma en su libro La Evolución de la Vida, [37] que «en lo que concierne al vuelo, hay algunas discontinuidades muy grandes en el registro fósil» (p. 180).
En lo que concierne a los insectos, dice Olson: «No hay prácticamente nada que nos informe sobre la historia del origen del vuelo en los insectos» (p. 180). En lo que trata de los reptiles voladores, Olson nos informa que «El vuelo verdadero está primeramente registrado entre los reptiles en las Pterosaurios del período Jurásico. Aunque los más primitivos de ellos estaban algo menos especializados para el vuelo que los últimos, no hay en absoluto ningún tipo o señal de etapas intermedias» (p. 181). Referente a las aves, Olson nos informa que «El vuelo verdadero está primeramente registrado entre los reptiles en los Pterosaurios del período Jurásico. Aunque los más primitivos de ellos estaban algo menos especializados para el vuelo que los últimos, no hay en absoluto ningún tipo o señal de etapas intermedias» (p. 181). Referente a las aves, Olson habla del Archaeopteryx como «reptiloide», pero afirma que por su posesión de plumas «muestra que es un ave» (p. 182). Finalmente, con referencia a los mamíferos, Olson afirma que «La primera evidencia de vuelo en los mamíferos es en los murciélagos ya completamente desarrollados del Eoceno» (p. 182 ).
El caso del Archaeopteryx: *
Así, pues, no se puede documentar ninguna serie de transición en ningún momento de la pretendida transición al vuelo y, tan sólo en un caso, se ha alegado la presencia de una forma de transición. En este último caso, el así llamado intermedio no es intermedio en absoluto, porque, tal y como los paleontólogos reconocen hoy en día, el Archaeopteryx era un ave verdadera –poseía alas, estaba dotada completamente de plumas, volaba. No era medio-ave. Era un ave. Jamás se ha encontrado una forma de transición con sólo parte de plumas y sólo parte de alas.
Las características que se alegan del Archaeopteryx consisten en los apéndices garróideos en los ejes-guía de las alas, la posesión de dientes, y vértebras que se extendían hacia afuera, formando una cola. Se cree qu e, poseyendo una quilla pequeña, era un volador más bien torpe. Mientras que tales características podrían ser de esperar si las aves evolucionaron de los reptiles, no constituyen por sí mismas una prueba en absoluto de que así haya sucedido, ni demuestran que el Archaeopteryx sea un intermedio entre los reptiles y las aves. Por ejemplo, en Sudamérica existe hoy en día un ave, el Hoatzin (Opisthocomus Hoatzin), el cual posee dos garras en su etapa juvenil. Aun más, es un volador torpe, con una quilla asombrosamente pequeña. [38] (36). Esta ave es –sin duda alguna– cien por cien ave, aun poseyendo dos de las características por las cuales se le imputa una ascendencia reptiliana al Archaeopteryx.
Mientras que las aves modernas no poseen dientes, otras aves más antiguas, aves cien por cien sin sombra de duda, los poseían. ¿Demuestra ello que los antepasados de las aves fueron los reptiles, o simplemente demuestra que algunas aves antiguas tenían dientes, mientras que otras no los tenían? Algunos reptiles tienen dientes y otros no los tienen. Algunos anfibios poseen dientes, otros no. De hecho, esto es cierto a través de todas las categorías de subphylums –peces, anfibios, reptiles, aves y mamíferos incluidos.
Siguiendo la analogía de que las aves dentadas son primitivas mientras que las desdentadas son más avanzadas, los monotremas (por ejemplo, el Ornitorrinco y el Equidna), ambos mamíferos, desdentados, ¡deberían ser considerados más avanzados que los humanos! Y a pesar de ello, estos mamíferos son considerados como los más «primitivos» de todos los mamíferos (Por cierto que se da la curiosa circunstancia de que ¡son los últimos en aparecer en el registro fósil!). ¿Qué valor filogenético se le puede dar a la posesión o no posesión de dientes?
En lo que se refiere a la posición del Archaeopteryx, DuNouy afirmó:
¿Cuál, pues, parece ser la conclusión más razonable? Creemos que el registro fósil no permite mejor secuela que la que Marshall ha publicado recientemente: «El origen de las aves es mayormente asunto de deducción. No hay fósiles de las etapas a través de las cuales se logró el asombroso cambia de reptil a ave». [40]
Objeciones
contestadas:
En vista de todo lo anterior, se presentan muchas excusas alegando que las estructuras delicadas generalmente no se preservan en forma fósil, para presentar una explicación al hecho de que no haya ninguna indicación, en el registro fósil, de la evolución de las plumas. Pero esta objeción no es posible en el caso de la pretendida evolución del vuelo en los reptiles voladores y los murciélagos.
Hay muchas diferencias, y muy significativas, entre reptiles voladores y no voladores. Otra vez nos remitimos a la obra de Romer, «Paleontología Vertebrada» (30). Se muestra en la página 140 la reproducción del Saltoposuchus (Fig. 214 de su libro), un representante de los Teocodontes del Triásico, que Romer cree dieron lugar a los reptiles voladores (Pterosaurios), dinosaurios y aves. La comparación de esta forma con las reconstrucciones de los más primitivos representantes entre los dos subórdenes de los Pterosaurios (pp. 144 y 146) revelan la vasta discontinuidad entre ellos, una discontinuidad no conectada por intermediarios fósiles. Desde luego, también existe una discontinuidad similar entre esta criatura y el Archaeopteryx.
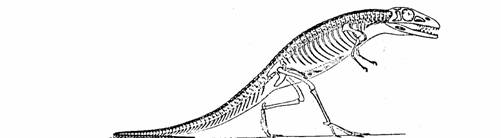
El
Saltoposuchus, teocodonte del que Romer y otros creen tuvieron su origen
los
dinosaurios, las aves, y los reptiles. Tomado de Paleontología Vertebrada, de
Romer.
Casi todas las
estructuras en el Rhamphorhyncus, pterosaurio de cola larga (Fig. 222, p. 144)
le eran peculiares. Es esencialmente obvia (como en todos los pterosaurios) la
gran longitud de su cuarto dedo, en contraste con los otros tres dedos que este
reptil poseía. Este cuarto dedo proveía todo el apoyo para la membrana que
actuaba de ala. Por cierto que no era una estructura delicada, y si los
pterosaurios, tuvieron su origen, en la evolución de los Tecodontes o de algún
otro reptil terrestre, se deberían haber hallado formas de transición mostrando
un alargamiento gradual de este cuarto dedo. No obstante, no se ha descubierto
ni una señal de tales formas de transición.
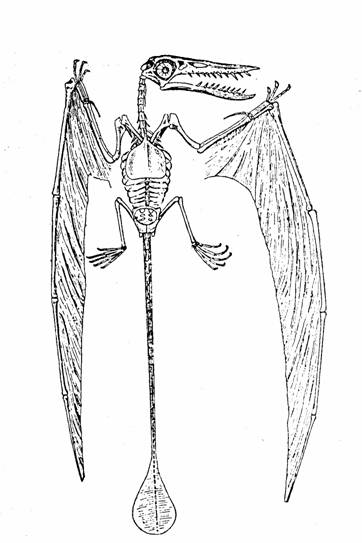
Rhamphorhyncus,
un Pterosaurio de cola larga.
Tomado de The Osteology of the Reptiles de Williston.
Aún más
especial era el grupo Pterodactiloide de los pterosaurios (Fig. 225, p. 146). El
Pteranodonte no solo tenía un pico largo y desdentado y un penacho óseo
largo extendiéndose hacia atrás, sino que sus cuatro dedos sostenían ¡un ala de
8 metros de envergadura! ¿Dónde están las formas de transición documentando el
origen evolutivo de estas y otras estructuras especiales de los Pterosaurios? En
cuanto al murciélago, se, supone que evolucionó de insectívoros terrestres, a
pesar de que, como se ha afirmado anteriormente, el murciélago más antiguo
conocido en el registro fósil [41] es cien por ciento
murciélago, y no se puede encontrar ni traza de formas de transición. En el
murciélago, cuatro de los cinco dedos sostienen la membrana del ala y son
extremadamente largos comparados con una mano normal. Estas y otras estructuras
únicas, como el radio y el cúbito, que se unen en la mitad distal para formar un
hueso simple en el Palaeochiropteryx (Romer, Fig. 322, p. 213), son huesos
sólidos, y no estructuras delicadas. Si las formas de transición existieron,
ciertamente que hubieran debido quedar preservadas. La completa ausencia de las
tales deja sin respuesta, tomando como base el modelo evolucionista, preguntas
tales como: ¿Cuándo, de qué, dónde y cómo tuvieron su origen los
murciélagos?
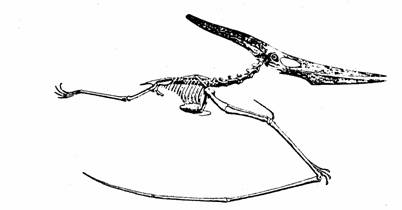
Pteronodonte, un
reptil gigante que tenía una longitud total de alas de más de
ocho metros.
Tomado de Paleontología Vertebrada
de Romer.
Ahora queremos
plantear la pregunta: En lo que concierne al origen del vuelo. ¿Qué modelo, el
de la creación, o el de la evolución, tiene mayor apoyo del registro fósil? A
nosotros la respuesta nos parece clara. Ni un solo hecho contradice las
predicciones del modelo creacionista: la verdadera evidencia falla en apoyar las
predicciones del modelo evolucionista. Aquí, donde sería más fácil y obvio el
hallar formas de transición si la evolución fuera realmente la causa de estas
criaturas tan altamente desarrolladas y especializadas, no se encuentra ni una.
¿Podría ser más cruel y caprichoso el registro fósil para los paleontólogos
evolucionistas? El registro histórico inscrito en las rocas está gritando
literalmente: ¡CREACIÓN!
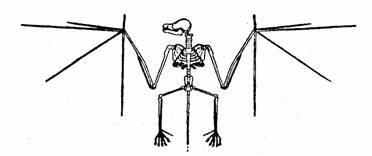
Palaeochiropteryx, el más antiguo de los murciélago
conocidos,
fósil del Eoceno.
Los ejemplos
citados en este artículo no son en ninguna manera excepciones, sino que sirven
para ilustrar lo que es característico del registro fósil. Mientras que se puede
observar una transición en el nivel de las subespecies, y se puede inferir en el
de las especies, la ausencia de las formas de transición entre las
categorías más elevadas de clasificación (los géneros creados del modelo
creacionista) es regular y sistemática. Nos proponemos documentar esta
afirmación citando afirmaciones públicas de evolucionistas.
AFIRMACIONES PUBLICAS DE
AUTORIDADES EVOLUCIONISTAS
(Referentes a la naturaleza del registro
fósil)
Deseamos citar primeramente al mundialmente famosísimo George Gaylord Simpson, paleontólogo evolucionista de primera fila. En su libro Tempo and Mode in Evolution (Ritmo y Método en la Evolución), afirma en la sección dedicada a «Discontinuidades Principales en el Registro (Fósil)» que no hay ni trazas en ninguna parte del mundo de ningún fó sil que cubra la discontinuidad que existe entre el Hyracotherium y su pretendida forma ancestral Condylarthra. Después sigue diciendo: «Esto es cierto de todos los treinta y dos órdenes de mamíferos... Los miembros más antiguos conocidos de cada orden ya tienen sus características ordinales básicas, y no hay, en ningún caso, una secuencia aproximadamente continua de un orden conocido a otro. En la mayoría de los casos, la rotura de conexión es tan brusca, y la discontinuidad tan grande, que el origen del orden es especulativo, y muy disputado». [42]
Después (p. 107), Simpson afirma:
«Esta ausencia regular de formas de transición no está limitada a los mamíferos, sino que es un fenómeno casi universal, y que ha sido notado desde hace mucho tiempo por los paleontólogos. Es cierto de casi todos los órdenes de todas las clases de animales, vertebrados e invertebrados. También es cierto de las clases, de los principales Phylums y, aparentemente, también es cierto de las categorías análogas de las plantas.»
En su libro The Meaning of Evolution (El Significado de la Evolución) Simpson afirma, refiriéndose a la aparición de nuevos phylums, clases y otros grupos principales:
«El proceso por el cual ocurrieron en la evolución unos sucesos tan radicales es causa de una disputa de las más serias que quedan entre los estudiosos profesionales de la evolución. La cuestión es si tales sucesos principales tuvieron lugar instantáneamente por algún proceso diferente en esencia de los que formarían parte en un cambio evolutivo más o menos gradual, o si toda la evolución, incluyendo estos cambios principales, queda explicada por los mismos procesos y principios directivos, siendo sus resultados mayores o menores en función del tiempo implicado, de la intensidad relativa de la selección y de otras variables en cualquier situación dada.
Si los phylums, clases, órdenes y otros grupos principales fueran conectados por formas de transición en l ugar de aparecer repentinamente en el registro fósil con sus características básicas completas, no habría necesidad, desde luego, de referirse a su aparición en el registro fósil como «sucesos radicales». Además ¡es imposible enfatizar con suficiente intensidad el que aun los mismísimos evolucionistas estén discutiendo entre ellos si estas categorías principales aparecieron instantáneamente o no! ¡Precisamente, el argumento de los creacionistas es que estas formas sí que se formaron instantáneamente, y que las formas de transición no están registradas en el registro fósil porque nunca existieron! Así, los creacionistas desearíamos reformar la afirmación de Simpson para que se leyese:
«Es así posible afirmar que tales transformaciones no están registradas porque no existieron, y que estos tipos principales se originaron por creación en lugar de por un proceso de evolución gradual.»
En un trabajo más reciente, Simpson afirmó que:«Es una característica del registro fósil el que la mayoría de los grupos taxonómicos aparecen bruscamente.»
En el mismo párrafo, además, afirma que:
A pesar de que es nuestro propósito hacerlo, apenas si es necesario documentar ya más la naturaleza del registro fósil. Parece obvio que si las afirmaciones anteriores de Simpson se desnudasen de todas las presuposiciones y de los supuestos mecanismos evolutivos para quedar simplemente el registro del hecho, describen exactamente lo que demanda el modelo creacionista. Este registro es, no obstante, terriblemente deficiente a la luz de las presuposiciones y predicciones del modelo evolucionista.
Nadie se ha entregado más de corazón que Simpson a la «Filosofía materialista mecanicista compartida por la mayor parte del Establecimiento actual en las ciencias biológicas. Así, como él mismo lo ha expresado, «Encuentra lógico, si no exigido científicamente, suponer que la aparición de un nuevo grupo sistemático no es evidencia de creación ...». [45]
Así, Simpson ha gastado un tiempo y un esfuerzo considerable en sus intentos de doblar y retorcer cada faceta de la teoría de la evolución para explicar las deficiencias del registro fósil, intentándolo acomodar a la hipótesis evolutiva. [46] , [47] , [48] Pero tenemos que hacer observar que, si se adopta la evolución a priori, siempre es posible adoptar hipótesis auxiliares –no probadas, y por su misma naturaleza imposibles de probar– para hacer que funcione en cualquier caso específico. Por este proceso, la evolución desemboca en lo que Thorpe llama uno de sus «Cuatro Pilares de la Ignorancia» –una evolución mental que es el resultado de tentativas casuales preservadas por apuntalamientos. [49]
Con referencia a la naturaleza del registro fósil, Arnold ha dicho:
Las siguientes afirmaciones del profesor E. J. H. Corner, de la Escuela Botánica de Cambridge (Universidad de Cambridge), fueron refrescantemente cándidas:
Olson ha dicho:
En relación con lo que algunos afirman sobre que el registro fósil esté (¡después de 150 años de búsqueda!) incompleto, nos remitimos a la afirmación de George ya citada al principio de este artículo sobre la gran riqueza del registro, y al hecho de que las discontinuidades, según el propio G. G. Simpson, son sistemáticas. Aun se puede citar a otro autor para otra refutación a esta superficial «explicación» de las discontinuidades en el registro fósil. Newell ha dicho:
En su reciente libro sobre los principios de la Paleontología, Raup y Stanley han afirmado:
DuNouy ha descrito la evidencia de esta manera:
«Brevemente, cada grupo, orden, o familia, parece haber nacido repentinamente, y a duras penas, si jamás, encontraremos las formas que las unen al grupo precedente. Cuando las descubrimos, están ya diferenciadas. No solamente no encontramos prácticamente ninguna forma de transición, sino que en general es imposible conectar un grupo nuevo con uno más antiguo». [55]
Khun ha enfatizado:
Tratando de los principales grupos, o Pylums, Clark ha afirmado:
Más adelante de este mismo volumen dice:
Aun en las famosas series equinas que tantas veces han sido alzadas en pendón como «prueba» de la evolución dentro de los órdenes, encontramos que las formas de transición entre los tipos principales faltan. Así, DuNouy ha afirmado en referencia a los caballos:
Goldschmidt dice en cuanto a esto mismo:
Goldschmidt, en contraste con Simpson y la mayoría de los evolucionistas, aceptó las discontinuidades en el registro fósil en su verdadero valor. Rechazó la interpretación Neodarwiniana de la evolución (la síntesis moderna en términos actuales de micromutación y Selección Natural) que es aceptada por casi todos los evolucionistas, por lo menos por aquellos que aceptan alguna teoría que implique algún mecanismo. Goldschmidt propuso, por su parte, que las categorías más importantes (Phylums, clases, órdenes, familias) se originaron instantáneamente por saltos bruscos o «systemic mutations» (mutaciones sistémicas). [61] , [62]
Goldschmidt bautizó a su mecanismo con el nombre de «Hopeful Monster» (Monstruo Viable). Propuso, por ejemplo, que un día un reptil puso un huevo, y que lo que salió del huevo fue ¡un ave! Según Goldschmidt, esta es la manera en que han de ser explicadas las principales discontinuidades del registro fósil, por sucesos similares. Algo puso un huevo, ¡y algo diferente nació! Los Neodarwinistas prefieren creer que el que puso el huevo es Goldschmidt, manteniendo que no hay ninguna evidencia que apoye el mecanismo del Monstruo Viable. Goldschmidt arguye con la misma intensidad en que no hay ninguna evidencia para el mecanismo Neodarwinista que se postula (grandes transformaciones por acumulación de micromutaciones). Los creacionistas estamos de acuerdo tanto con Goldschmidt como con los Neodarwinistas: ¡Ambos están completamente equivocados! Las publicaciones de Goldschmidt ofrecen argumentos convincentes en contra del punto de vista Neodarwinista de la evolución, desde el campo de la genética y de la paleontología.
Nadie estaba más verdaderamente entregado a la filosofía evolucionista que Goldschmidt. Si alguien deseaba encontrar formas de transición, él más. Si alguien hubiera tenido que admitir que una forma de transición lo fuera, él hubiera sido el primero. Pero en cuanto al registro fósil, he aquí lo que Goldschmidt tuvo que decir:
Ahora preguntamos: ¿Qué mejor descripción del registro fósil se podría esperar, en base de las predicciones del modelo de la creación? Por otra parte, a no ser que se acepte el mecanismo evolutivo de Goldschmidt –el «Monstruo Viable»– esta descripción contradice la predicción más crítica del modelo de la evolución: la presencia de los intermediarios exigidos por la teoría en el registro fósil.
Kerkut, a pesar de no ser un creacionista, escribió un pequeño pero notable volumen para exponer la debilidad y las falacias en la evidencia generalmente utilizada para apoyar la teoría de la evolución. En la fase final de su libro Kerkut afirmó que:
Hay mundos de diferencia, desde luego, entre una hipótesis de trabajo y un hecho científico establecido. Si las presuposiciones filosóficas le guían a uno a aceptar la evolución como su hipótesis de trabajo, la debería restringir a este uso, y no obligar a su aceptación como hecho establecido.
Sí, dejando aparte cualquier presuposición filosófica, sea materialista, o teísta, se utilizan la creación y la evolución como modelos para predecir la naturaleza de la evidencia histórica que se espera ha de coincidir con uno de ellos, podemos ver que el modelo de la creación es tan creíble (y nosotros creemos que mucho más) como el modelo de la evolución. Reiteramos que ningún modelo es más religioso ni menos científico que el otro.
Nadie menos que un evolucionista convencido como Thomas H. Huxley reconoció que:
La mayoría en la comunidad científica y en los círculos educativos
están usando la excusa de la «ciencia» para forzar la enseñanza de su punto de
vista sobre la vida sobre todo el mundo. El autoritarismo de la Iglesia Medieval
ha sido reemplazado por el autoritarismo del Materialismo Racionalista. Se viola
el respeto mutuo, y la libre investigación queda ahogada bajo esta capa de
dogmatismo. Es hora ya de cambiar.
(*) Este
artículo fue leído ante la Convención Anual de la Asociación Nacional de
Profesores de Biología en San Francisco (California). Volver al texto
 Ir a la
siguiente sección del libro
Ir a la
siguiente sección del libro
Volver al Índice de Creación, Evolución y el Registro
Fósil
Volver
a libros Creación y Ciencia
Volver a la página principal
Volver al índice general
castellano
Agradecimientos
SEDIN
(Servicio Evangélico de Documentación e Información ) desea manifestar su
profundo agradecimiento a las siguientes entidades y personas que han hecho
posible la presentación en castellano de esta serie Creación y Ciencia,
facilitándonos los permisos para la publicación del material que aquí
presentamos:
INSTITUTE FOR
CREATION RESEARCH, de San Diego, California, dirigido por los Drs. Henry M.
Morris y Duane T. Gish.
CREATION RESEARCH SOCIETY, de Ann Arbor, Michigan, y el Managing Editor, Dr. John N. Moore.
BIBLE-SCIENCE ASSOCIATION, de Minneapolis, Minnessota, dirigida por el Rev. Walter Lang.
Dr. JOHN C. WHITCOMB, Profesor de Antiguo Testamento en el Grace Theological Seminary de Winona Lake, Indiana, y a la editorial Presbyterian & Reformed Pub. Co.
Mr. R. G. ELMENDORF, ingeniero de Bairdford, Pennsylvania.
SPIRITUAL COUNTERFEITS PROJECT, y la BERKELEY CHRISTIAN COALITION de Berkeley, California.
SEDIN
Apdo. 2002
08200 SABADELL
(Barcelona) ESPAÑA
LIBROS CREACIÓN y
CIENCIA
CREACIÓN,
EVOLUCIÓN Y EL REGISTRO FÓSIL
© Institute for
Creation Research; Creation Research Society; Presbyterian and Reformed
Publishing Co.
© 1977 SEDIN, Apartado 2002 - 08200 SABADELL (Barcelona), ESPAÑA para la versión española
© 2003 SEDIN, Apartado 2002 - 08200 SABADELL (Barcelona), ESPAÑA para la versión electrónica
Compilado por
SEDIN, con permiso de Institute for Creation Research; Creation Research
Society; Presbyterian and Reformed Publishing Co.
- Traducción del inglés: Santiago Escuain
© Santiago Escuain 1977, por la traducción
© Copyright SEDIN 2003 para el formato electrónico - www.sedin.org. Este texto se puede reproducir libremente para fines no comerciales y citando la procedencia y dirección de SEDIN, así como esta nota en su integridad.
SEDIN
Servicio Evangélico - Documentación -
Información
Apartado 2002
08200 SABADELL
(Barcelona) ESPAÑA
Índice:
Índice de
boletines
Índice
de línea
sobre línea
Página
principal
Índice
general castellano
Libros recomendados
orígenes
vida
cristiana
bibliografía
general
Coordinadora
Creacionista
Museo de
Máquinas Moleculares
Temas de
actualidad
Documentos en
PDF
(clasificados por temas)




||| General English Index ||| Coordinadora Creacionista ||| Museo de Máquinas Moleculares ||| ||| Libros recomendados ||| orígenes ||| vida cristiana ||| bibliografía general ||| ||| Temas de actualidad ||| Documentos en PDF (clasificados por temas) ||| |